Die Regulation des Kreislaufs
Die Informationen auf dieser Seite sind auch in einem eBook der Patienten-Akademie enthalten.
Sie bekommen die Version dieses eBooks (Band 4 der eBook-Reihe) in verschiedenen Formaten:
- padBook (für iPad und epub3-fähige eBook-Reader)
- phoneBook (für smartPhones)
- Paperwhite (für Kindle Paperwhite)
Vorbemerkungen
Die Funktion des Kreislaufs ist für das Überleben eines Organismus von grundlegender Bedeutung, denn es ist der Kreislauf, der alle Organe mit lebenswichtigem Sauerstoff und Nährstoffen versorgt. Arbeitet er nicht optimal kommt es zu Störungen der Organfunktionen, bricht er zusammen stirbt das Lebewesen.
Nun reicht es nicht aus, wenn man die Funktionsgrößen des Kreislaufs, also z.B. Höhe des Blutdrucks oder Menge des vom Herzen ausgepumpten Blutes, einmal fest einstellt, den Kreislauf dann startet und ihn einfach mit diesen fest eingestellten Größen laufen läßt. Der Kreislauf ist nämlich von einer Vielzahl von Einflüssen abhängig, die seine Arbeitsweise beeinflussen, z.B. Wassermangel ebenso wie ein Überschuß an Wasser im Kreislauf, Sauerstoffmangel, Hitze, Kälte, Fieber, Aufenthalt in großer Höhe oder Ähnliches.
Solche Faktoren können zu einer u.U. erheblichen Störung des Kreislaufes führen, die die permanente Blutversorgung der Organe beeinträchtigen oder sogar gefährden. Daher muß der Organismus in der Lage sein, die Auswirkungen solcher Störungen zu erkennen und zu korrigieren.
Und genau mit diesem Aspekt des Kreislaufes, d.i. mit seiner Regulierung, möchte ich mich in diesem eBook geschäftigen.
Die Regulierung des Kreislaufes ist kompliziert, weil hier verschiedene Akteure beteiligt sind, die in unterschiedlicher Weise mit- und zusammenarbeiten. Ich kann Ihnen dies an einem hoffentlich gut verständlichen Beispiel zeigen, nämlich dem Brand in einem Bürogebäude:
- In einer kleinen Kaffeeküche gibt es einen Kurzschluß in der Kaffeemaschine. Die Maschine beginnt zu brennen und Rauch steigt auf.
- Der Rauchmelder in der Küche bemerkt den Rauch und schlägt mit schrillem Warnton an.
Es geschehen nun 3 Dinge:
- Der Rauchmelder löst automatisch die Sprinkleranlage aus.
- Es wird die Feuerwehr alarmiert, und zwar sowohl die benachbarte (örtliche) Feuerwache als auch die Hauptfeuerwache.
- Die Sprinkleranlage verspritzt Wasser, mit dem die brennende Kaffeemaschine sofort und schnell gelöscht werden soll.
- Die örtliche Feuerwache alarmiert den hier stationierten Löschzug über die hausinterne Alarmleitung.
- Die Besatzung der Feuerwehr-Hauptwache versammelt sich im Kontrollraum und beobachtet die Signale des Rauchmelders aus Ihrer Wohnung.
- Der Löschzug kommt an und sieht sich das Problem an: Haben die Sprinkler das Feuer gelöscht oder brennt es noch?
- Wenn das Feuer bereits gelöscht ist:
- Der Löschzug informiert seine (örtliche) Feuerwache und rückt wieder ab.
- Die Rauchmelder in der Küche geben nun keinen Alarm mehr. Feuer- und Hauptfeuerwache können nun davon ausgehen, daß das (Brand-) Problem erfolgreich beseitigt wurde. Alle entspannen sich und setzen ihre üblichen Arbeiten fort
- Der gesamte Alarm wird beendet.
- Wenn das Feuer nicht gelöscht ist:
- Der Löschzug beginnt mit eigenen Löscharbeiten.
- Die örtliche Feuerwache steht mit dem Löschzug in Verbindung und entscheidet darüber, ob ggfs. ein weiterer Löschzug geschickt werden muß.
III. Auch die Hauptfeuerwache wird den erfolglosen 1. Löschversuch zur Kenntnis nehmen, weil nämlich die Signale des Rauchmelders nicht verstummen. Sie wird nun darüber zu entscheiden haben, was weiter zu tun ist, um das bestehende Problem zu beseitigen und sie wird die entsprechenden Maßnahmen in die Wege leiten, z.B.:
- Neuen Kaffee kochen, weil das alles kann jetzt dauern
- Feuerwehren umliegender Städte informieren
- Löschhubschrauber anfordern
- die Polizei informieren (um das Brandgebiet abzusperren)
- den Notarzt informieren (um evtl. Verletzte zu versorgen)
- die Stadtwerke informieren (um den Strom im brennenden Haus abzustellen). Diese ganzen Maßnahmen werden erst dann beendet, wenn der Rauchmelders am Brandherd keine weiteren Signale mehr gibt (Es handelt sich um einen Neubau, in dem Sie wohnen, daher ist der Rauchmelder davor geschützt, in den Flammen zu schmelzen, dadurch zerstört zu werden und allein aus diesem Grund keinen Alarm mehr geben zu können).
Wenn Sie sich diese Kette von Ereignissen und Maßnahmen genau ansehen werden Sie verschiedene Dinge und Akteure erkennen, die für die Lösung des Problems notwendig sind:
- Rauchmelder: Er arbeitet als Sensor und ist dazu geschaffen, Rauch und damit ein Problem zu erkennen, dieses Problem zu melden und gleichzeitig 1. Gegenmaßnahmen einzuleiten.
- Das Wasser, das die Sprinkler und der Schaum, den der Löschzug versprühen sind die Werkzeuge zur Lösung des Problems.
- Der Schlauch, mit dem der Löschzug den Schaum ins Feuer sprüht ist ein Transport für das Werkzeug.
- Örtliche Feuerwache und Hauptfeuerwache sind die Steuerzentralen, die dafür zuständig sind, die Lösung des Problems zu organisieren und zu leiten.
- Der Sensor alarmiert die Feuerwachen über Telephon oder Funk, ebenso wie die Kommunikation zwischen beiden Wachen und dem Löschzug über Telephon oder Funk erfolgt. Hierbei handelt es sich um die Kommunikationsleitung.
Alle diese Komponenten der Kreislaufregulierung, also Sensoren, Werkzeuge, Transportmittel für die Werkzeuge, Steuerzentralen und Kommunikationswege, sorgen dafür, daß immer der Normalzustand herrscht, also daß z.B. der Blutdruck weder zu hoch noch zu tief ist. Zu diesem Zweck müssen alle Komponenten miteinander verbunden sein, sodaß die Intensität evtl. Gegenmaßnahmen durch die Werkzeuge von den Informationen der Sensoren gesteuert werden. Dies erfolgt im Rahmen von sog. Regelkreisen, über die ich Ihnen später in diesem eBook noch genauer berichten werde.
Zunächst möchte ich Sie jedoch mit Sensoren, Werkzeugen, Transportmitteln für die Werkzeuge, mit Kommunikationswegen und Steuerzentralen bekannt machen.
Sensoren
Sie arbeiten als „Meßfühler“ und dienen dazu, bestimmte „Meßgrößen“ (oder auch „Parameter“) zu erfassen.
Solche Parameter sind z.B.
- Blutdruck,
- Wandspannung und Wanddehnung von Gefäßen und Herzmuskel,
- Säuregehalt des arteriellen und venösen Blutes (pH-Wert),
- Sauerstoffgehalt der Atmungsluft und in den verschiedenen Geweben,
- Gehalt des Blutes, der Gewebes und des Urins an verschiedenen Salzen (z.B. Natrium, Kalium),
- Temperatur des Gewebes
- Gehalt des Gewebes an ADP.
Gemeinsam ist allen diesen Sensoren, daß sie solche Vorgänge im Körper und im Stoffwechsel der Zellen messen, die für die normale Funktion des Organismus von ausschlaggebender Bedeutung sind und die unbedingt in Ordnung gehalten werden müssen, weil anderenfalls der Zusammenbruch des gesamten System drohen würde.
Bevor ich einige spezielle Sensoren vorstelle möchte ich Ihnen zunächst etwas Allgemeines über Sensoren und deren Arbeitsweise erklären:
Zunächst einmal handelt es sich bei den Sensoren des Menschen (ebenso wie diejenigen meines Dackels Heidi) nicht um technische Geräte wie etwa ein Fieberthermometer oder ein Meßgerät, mit denen man den Druck in einem Autoreifen mißt. Man spricht in der Biologie vielmehr von „Rezeptoren“.
Bauprinzip einer Rezeptorzelle
 |
| Abb. 1 |
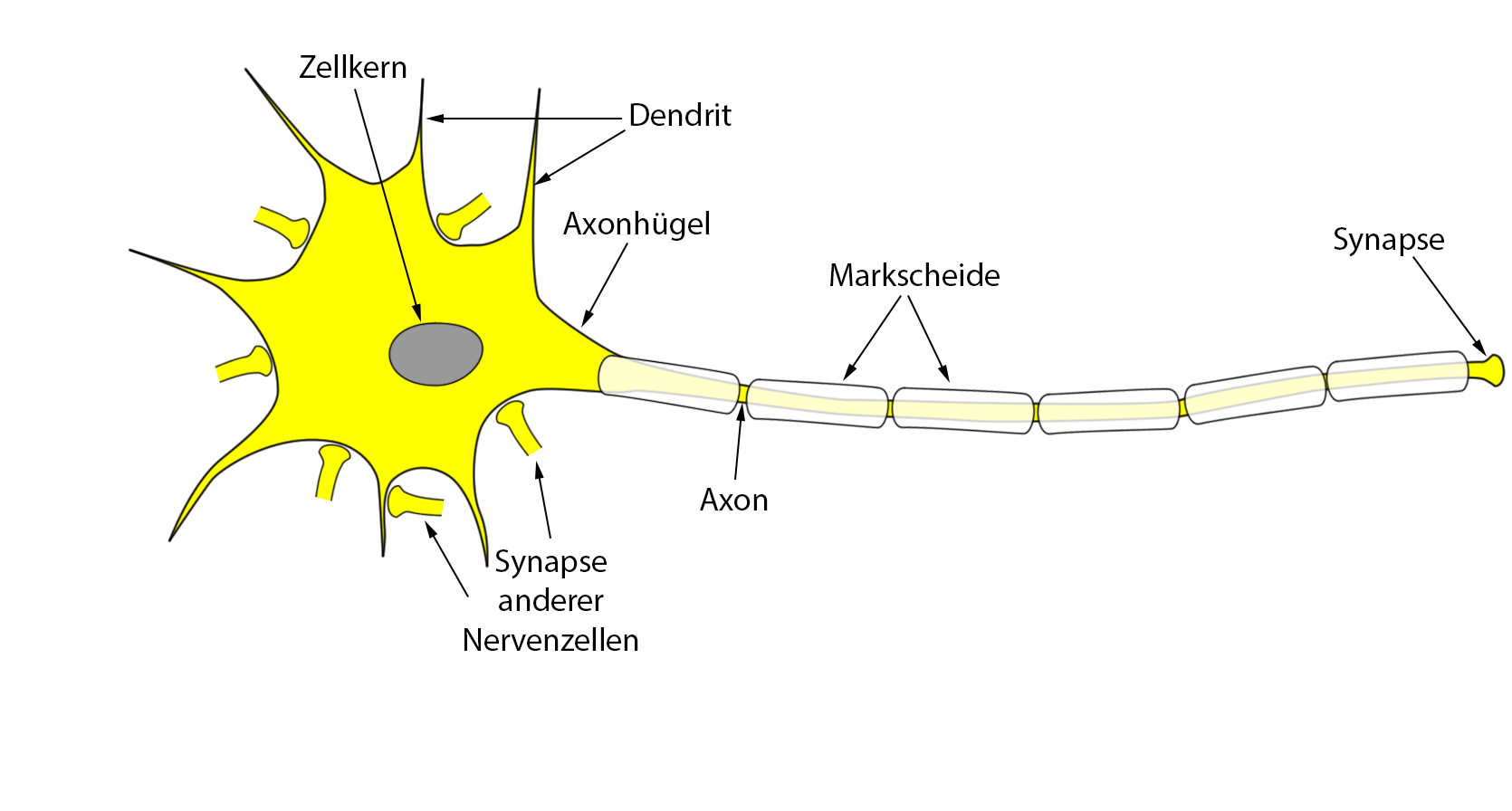
Rezeptoren sind (bis auf wenige Ausnahmen) spezialisierte Nervenzellen und daher bestehen sie, wie alle anderen Nervenzelltypen auch (Abb. 1), aus einem
- Zellkörper mit einem Zellkern und allen anderen Strukturen, die eine Zelle benötigt (z.B. Mitochondrien zur Energiegewinnung) und
- vom Zellkörpers abgehenden Ausstülpungen.
Dabei gibt es kurze (Dendriten) und lange (Axone) Ausstülpungen.
Art und Funktion dieser Ausstülpungen sind es, die eine Rezeptorzelle von einer „normalen“ Nervenzelle unterscheiden.
Normale Nervenzellen dienen der Leitung und Übertragung elektrischer Impulse (sog. Erregungen) von einer Nerven- zu einer anderen Nervenzelle oder von der Nervenzelle auf ein „Zielorgan“, z.B. einen Muskel, auf Drüsen oder die Muskelwände von Blutgefäßen.
| Abb. 2 |
Die kurzen Dendriten der normalen Nervenzelle dienen dazu, Impulse von anderen Nervenzellen zu empfangen, die Axone dazu, Verbindung an andere Nervenzellen zu übertragen (Abb. 2).
 |
| Abb. 3 |
Rezeptorzellen sind etwas anders gebaut:
Sie haben ebenfalls Dendriten und ein Axon, die aber miteinander verschmolzen sind und sich kurz nach ihrem Ursprung wieder teilen und zwar in ein Axon und einen Axon-ähnlichen Dendriten. Man nennt solche Zellen „pseudounipolare Nervenzellen“ Abb. 3).
Am Ende des Axon-ähnlichen Dendriten befindet sich bei der Rezeptorzelle der eigentliche Rezeptor.
Er mißt die „Meßgröße“, für die er spezifisch geschaffen wurde (z.B. Blutdruck, Sauerstoffgehalt oder Temperatur).
Funktionsprinzip eines Rezeptors
Mit dem Begriff „Rezeptor“ (oder auch Sensor bzw. Sinneszelle genannt) bezeichnet man spezialisierte Zellen, die in der Lage sind, bestimmte physikalische (z.B. Blutdruck, Dehnung eines Gewebes), chemische Zustände (z.B. Säuregehalt des Blutes, Sauerstoffgehalt bestimmter Gewebe) oder die Temperatur wahrzunehmen. Dies erfolgt dadurch, daß bestimmte Vorgänge im Körper zu chemischen oder molekularen Veränderungen im Rezeptor führen.
Diese chemischen oder molekularen Veränderungen wiederum führen zur Ausbildung elektrischer Impulse in den Zellen.
 |
| Abb. 4 |
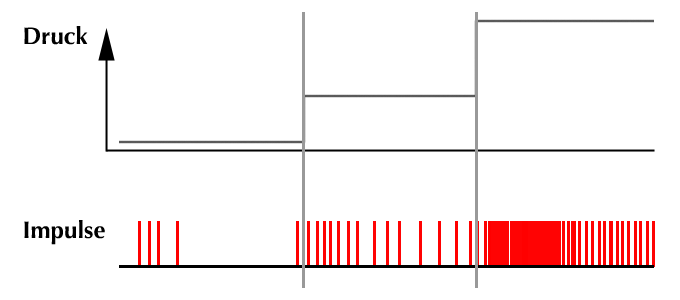
Die Art dieser Impulse, also ihre Häufigkeit (= Frequenz) und ihre Stärke werden von der „Intensität“ des Parameters beeinflußt: Befindet sich der Parameter im „Normalzustand“ werden Impulse einer bestimmten Frequenz produziert. Ist der Parameter höher als im Normalzustand werden mehr, ist er niedriger als der Normalzustand werden weniger Impulse gebildet.
Sehen Sie dies beispielsweise in Abb. 4 für einen Blutdruckrezeptor:
Ist der Blutdruck höher als normal (rechter Teil der Abbildung) werden mehr Impulse gebildet als bei normalem (Mitte des Bildes) oder niedrigem Blutdruck (linker Teil des Bildes).
Die Frequenz der Rezeptorimpulse läßt also Rückschlüsse auf die gemessene Höhe des jeweiligen Parameters zu.
Rezeptor-Typen
Bei den meisten Rezeptoren werden diese Informationen nun zu bestimmten Steuerungszentren weitergeleitet, die auf der Basis dieser Informationen Entscheidungen fällen, wie weiter zu verfahren ist. Diese Rezeptoren haben eine ausschließlich messende und informierende Funktion, ich würde sie daher als „informative Rezeptoren“ bezeichnen.
Es gibt aber auch andere Rezeptoren, die Veränderungen der von ihnen wahrgenommenen Meßgröße nicht nur messen, sondern die gleichzeitig auch bestimmte chemische Substanzen (z.B. Hormone) produzieren. Dies trifft beispielsweise auf die Dehnungsrezeptoren in den Wänden der Vor- und Hauptkammern des Herzens. Einerseits senden sie ihre Impulse an bestimmte Steuerungszentren, andererseits bilden sie bestimmte Hormone, nämlich „ANP“ und „BNP“, über die Sie später noch hören werden.
Solche Rezeptoren, die sowohl als Sensor als hormonproduzierendes Werkzeug arbeiten nenne ich „kombinierte Rezeptoren“.
Und dann gibt es schließlich noch solche Rezeptoren, die keine Impulse an übergeordnete Steuerungszentren schicken, sondern die in Abhängigkeit von ihren Messungen bestimmte Substanzen produzieren und ins Blut ausschütten. Ein Beispiel hierfür ist der juxtaglomerulären Apparat in der Niere, der in bestimmten Situationen das Hormon „Renin“ bilden, das in den Wasserhaushalt des Körpers eingreift (Sie werden später Genaueres erfahren). Solche Rezeptoren würde ich als „hormonproduzierende Rezeptoren“ bezeichnen.
Informationsübertragung der Rezeptoren
Informative und kombinierte Rezeptoren sind, wie schon erwähnt, spezialisierte Nervenzellen. Sie haben also, wie das bei Nervenzellen üblich ist, die Möglichkeit, elektrische Impulse zu leiten:
 |
| Abb. 5 |
Vom eigentlichen Rezeptor an der Spitze des Dendriten aus werden die Impulse zunächst in Richtung auf den Nervenzellkörper und von hier aus über das Axon bis zu dessen Ende fortgeleitet. Einige Axone und Dendriten sowohl einer „normalen“ Nerven- als auch von Rezeptorzellen sind von einer sog. „Markscheide“ aus speziellen Eiweißen umgeben (Abb. 5).
Sie sollen das Axon bzw. den Dendriten gegenüber seiner Umgebung isolieren. Bei anderen Axonen bzw. Dendriten fehlt eine solche Markscheide.
Axone bzw. Dendriten mit der Markscheide leiten elektrische Impulse wesentlich schneller als solche ohne eine Scheide.
Am Ende des Axons erfolgt die Übertragung der Impulse auf eine andere Nervenzelle. Diese Verbindungs- und Übertragungsstelle nennt man „Synapse“.
Synapse
Synapsen sind diejenigen Stellen, über die eine Nervenzelle in Kontakt zu anderen Nervenzellen steht.
Dies betrifft auch die Rezeptorzellen, die ja auch Verbindung zu anderen Nervenzellen aufnehmen müssen, damit ihre Informationen (d.i. ihre Impulse) weitergeleitet werden können.
 |
| Abb. 6 |
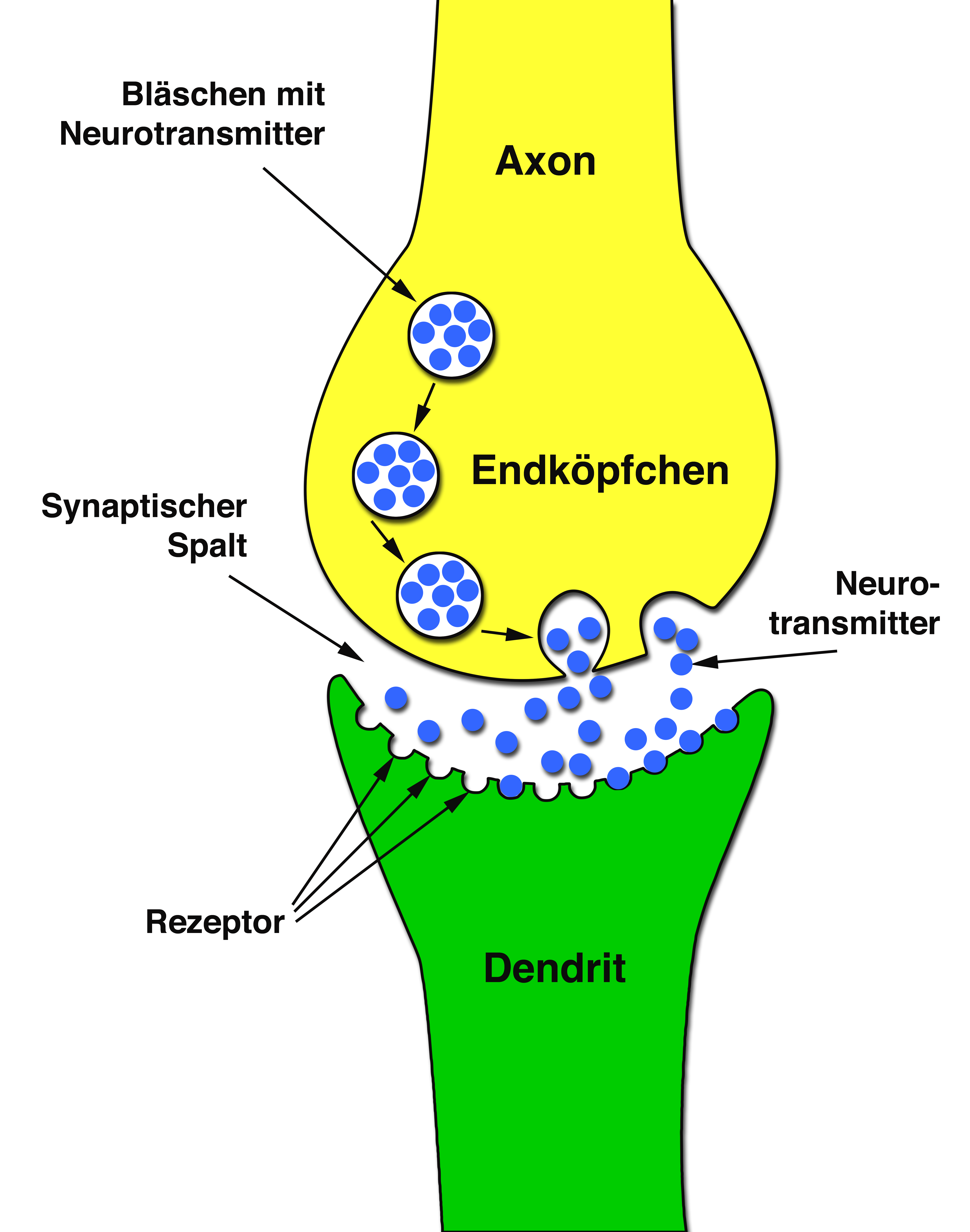
Beteiligt sind am Aufbau einer Synapse das Endköpfchen des Axon und der Dendrit einer anderen (Empfänger-) Zelle (Abb. 6).
Im Endköpfchen einer Synapse befinden sich winzige Bläschen, in denen sich sog. Botenstoffe (Neurotransmitter) befinden. Bei diesen Neurotransmittern handelt es sich (in Abhängigkeit vom Nerventyp und der Art der Synapse) um unterschiedliche chemische Substanzen, z.B. Acethylcholin, Adrenalin, Noradrenalin, Dopamin oder gamma-Aminobuttersäure.
Das Bläschen selber reagiert auf Calcium-Ionen:
Gelangt ein elektrischer Impuls über das Axon zu dessen Endköpfchen verursacht es hier den Einstrom von Calcium.
| Film 1 |
Dieser Calcium-Einstrom veranlaßt das Bläschen, mit der Membran des Endköpfchens zu verschmelzen. Dabei entleert sich der Inhalt des Bläschens, d.i. der Botenstoff in den Raum zwischen Endköpfchen und Dendrit der angeschlossenen benachbarten Zelle (synaptischer Spalt) (Film 1).
Auf der Oberfläche des Dendriten wiederum befinden sich sog. „Rezeptoren“, die nun in Kontakt mit dem Botenstoff kommen. („Rezeptoren“ sind in diesem Zusammenhang Eiweiße, die in die Membran von Zellen eingebaut sind und die wegen ihrer speziellen Form nur auf bestimmte andere Moleküle reagieren, die sich ihnen anlagern können.)
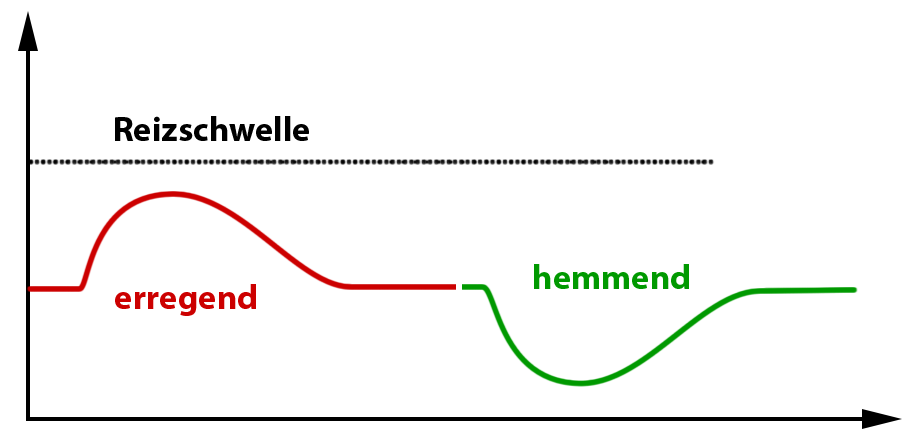
Im Fall einer Nervenzellen-Synapse gibt es 2 Typen von Rezeptoren, einen erregenden und einen hemmenden, die jeweils auf spezielle Neurotransmitter reagieren:
 |
| Abb. 7 |
- Acetylcholin beispielsweise reagiert mit dem erregenden Rezeptor. Dies hat zur Folge, daß sich Membrankanäle öffnen, die für positiv geladene Ionen (z.B. Natrium) durchgängig sind. Strömen nun viele positiv geladene Ionen in das negativ geladene Innere der Nervenzelle nimmt das Membranpotential ab (Abb. 7) und erreicht, wenn genügend Rezeptoren aktiviert wurden, die Reizschwelle. Hierdurch wird ein Aktionspotential, d.h. ein elektrischer Impuls ausgelöst, der dann über die gesamte Nervenzelle weitergeleitet wird. An der Synapse wird somit der elektrische Impuls von einer auf eine andere Nervenzelle übertragen, sodaß man in diesem Fall von einem erregenden Synapse bzw. einer „erregenden Reaktion“ spricht.
- Ein anderer Rezeptortyp reagiert nicht auf Acetylcholin, sondern mit gamma-Aminobuttersäure. Bei diesem Rezeptortyp löst der Botenstoff die Öffnung von Chlorid-Kanälen aus. Chlorid ist ein negativ geladenes Ion. Gelangen nun viele negativ geladene Ionen in das ohnehin schon negativ geladene Zellinnere dann nimmt das Membranpotential weiter zu (Abb. 7) und entfernt sich weiter von der Reizschwelle. In diesem Fall kann natürlich kein Aktionspotential ausgelöst werden, d.h. daß an dieser Synapse kein Impuls übertragen wird. Man spricht daher von von einer hemmenden Synapse bzw. einer hemmenden Reaktion.
Auf diese Weise werden Informationen entweder von der Nervenzelle auf eine andere Zelle weitergegeben oder die Weitergabe der Impulse wird blockiert.
Für die Arbeitsweise der verschiedenen Regulationsmechanismen ist dies von großer Bedeutung, wie Sie später noch lesen werden.
Ein weiterer wichtiger Begriff im Zusammenhang mit Nerven- und Rezeptorzellen ist die Richtung, in der die Impulse und damit die Informationen übertragen werden und der anatomische Ort, an dem sich die Körper der Rezeptorzellen befinden.
Richtung der Informationsleitung
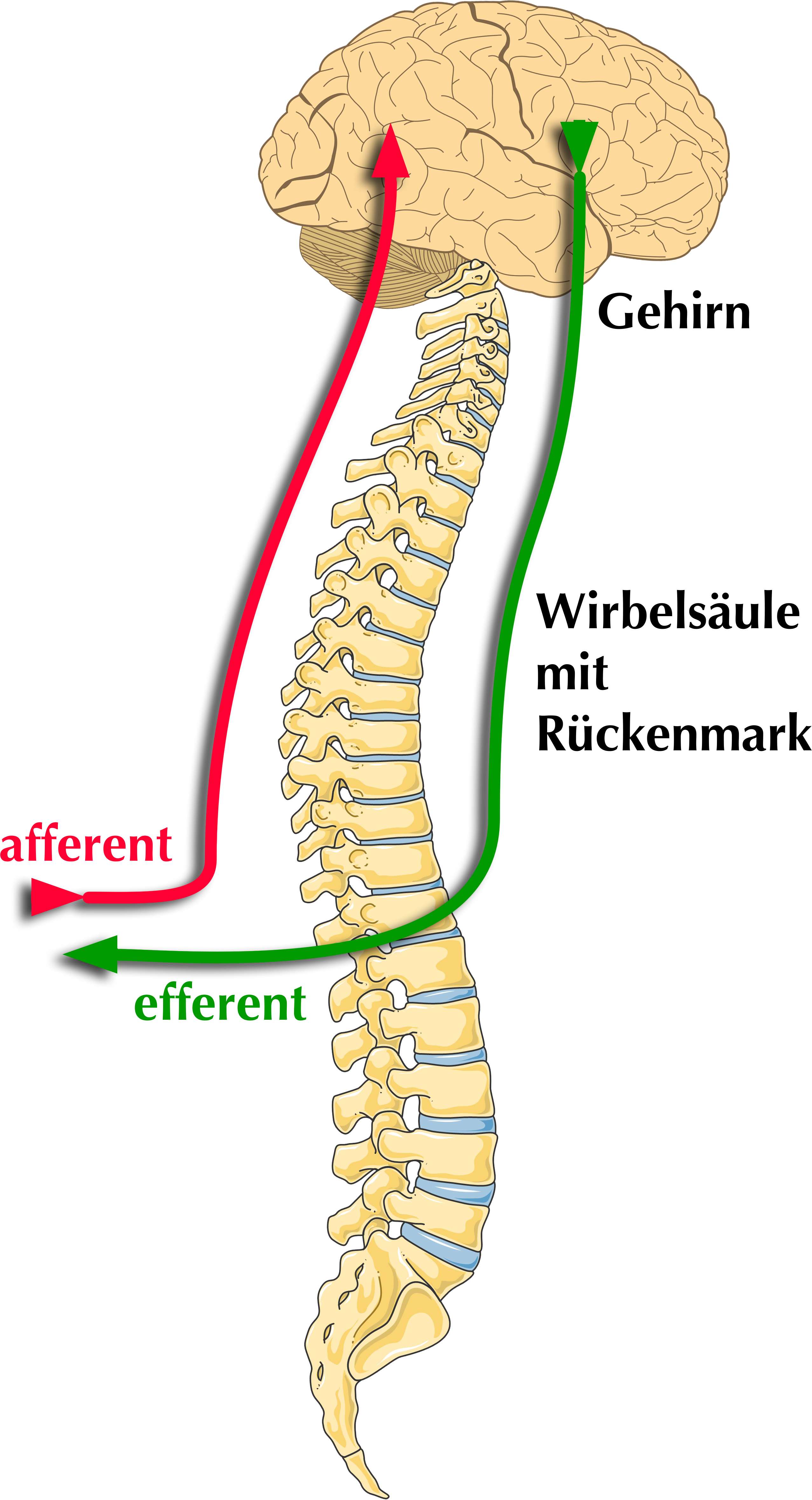 |
| Abb. 8 |
In Bezug auf die Richtung innerhalb des Körpers, in die Nerven ihre Impulse, d.h. ihre Informationen übertragen unterscheidet man afferente und efferente Nerven (Abb. 8).
Afferente Nerven leiten ihre Impulse zum Zellkörper der Nerven- oder Rezeptorzelle hin, efferente Nerven führen Impulse vom Zellkörper weg zu Muskeln, Drüsen oder anderen Nervenzellen.
In diesem Zusammenhang spricht man auch von afferenten und efferenten Impulsen:
Afferente Impulse führen auf den Zellkörper zu, efferente Impulse laufen vom Zellkörper weg. Sie werden diese Begriffe im Folgenden häufiger hören.
Lokalisation des Zellkörpers
Es ist für das Verständnis der Funktion und des Aufbaus eines Rezeptors wichtig zu wissen, wo sich sein „Ursprung“, d.h. der Zellkörper befindet, denn von diesem Ursprung aus breiten sich Axon und Dendrit aus.
Die Zellkörper der Rezeptoren sind nicht alle an einem Ort versammelt. Wo sie sich befinden hängt im wesentlichen von der Art des Rezeptors ab, ich werde dies etwas später unter den einzelnen Rezeptoren beschreiben.
Und nun wie angekündigt zu einigen wichtigen und elementaren Rezeptoren:
Die einzelnen Rezeptoren
Barorezeptor
Typ: Informativer Rezeptor
Es handelt sich um einen Sensor, der Druck „messen“ kann („Baro“ ist eine Bezeichnung für „Druck“; das Wort ist abgeleitet aus dem griechischen „baros“ für „Schwere“ und „Gewicht“).
Barorezeptoren gehören zur Gruppe der „Mechanorezeptoren“, sie sind rein informative Rezeptoren, d.h. sie übertragen lediglich das Ergebnis Ihrer Messungen ohne selber (z.B. durch die Produktion von Hormonen) tätig zu werden.
Man unterscheidet „Hoch-“ und „Niederdruck-Rezeptoren“:
- Die Hochdruckrezeptoren sind zuständig für die Messung des Blutdrucks in den Schlagadern. Sie sind es, die man als eigentliche „Druckrezeptoren“ bezeichnet.
- Die Niederdruckrezeptoren messen den u.a. Füllungszustand der Vorkammern des Herzens. Sie werden Dehnungsrezeptoren genannt und werden im nächsten Absatz besprochen.
Weil ihre Aufgabe die Messung des Blutdrucks ist befinden sich die Druckrezeptoren in den Wänden vieler Arterien, dort sind sie eher diffus über die Gefäßwände verteilt.
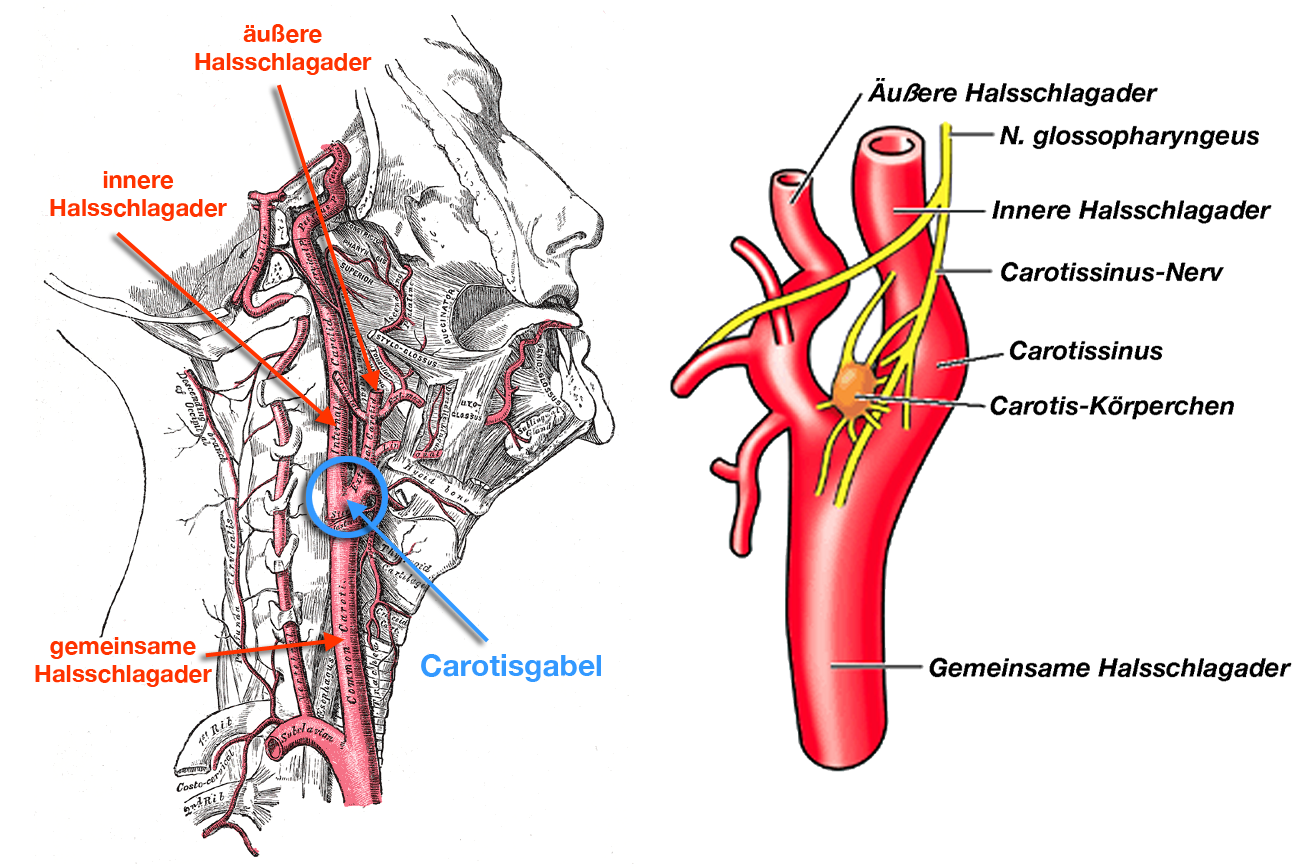 |
| Abb. 9 |
Es gibt aber Stellen in den Schlagadern, an denen sich zahlreiche solcher Druckrezeptoren örtlich konzentriert und geballt finden. Die prominentesten Beispiele hierfür sind die Aorta im Brustbereich und der sog. Carotissinus (Abb. 9), von dem etwas später noch die Rede sein wird.
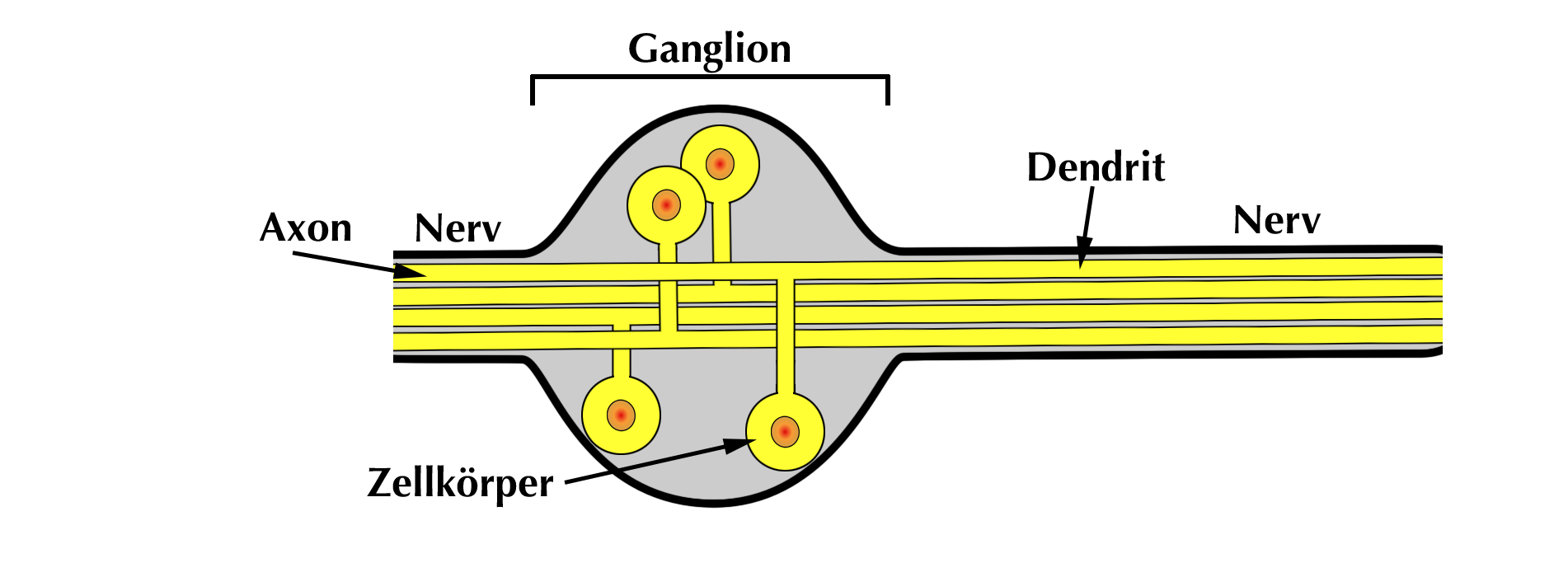 |
| Abb. 10 |
Druckrezeptoren sind spezialisierte Nervenzellen und daher sind sie ebenso aufgebaut wie diese.
Sie bestehen aus einem Nervenzellkörper mit seinen beiden Ausziehungen, dem Axon und dem Dendriten. Die Zellkörper liegen geballt in sog. Ganglien liegen (Abb. 10), über die ich etwas später genauer berichten werde.
Die Zellkörper derjenigen Barorezeptoren, die die Aortenwand versorgen liegen im Ganglion des 10. Hirnnerven (N. vagus). Man nennt es Ganglion nodosum und es liegt im Bereich der Schädelbasis.
Dicht daneben an der Schädelbasis liegt das sog Ganglion petrosum mit den Nervenzellkörper derjenigen Druckrezeptoren, die den Carotissinus der Halsschlagader versorgen. Dieses Ganglion gehört zum 9. Hirnnerven, dem N. glossopharyngeus.
Der Axon-ähnliche Dendrit (siehe Abb. 3) einer Rezeptorzelle trägt an seinem Ende den eigentlichen Drucksensor. Er ist so gebaut, daß er auf seine Dehnung reagiert. Die Grundüberlegung ist dabei, daß sich ein Blutgefäß, in dem ein hoher Blutdruck herrscht, rein mechanisch erweitert und daß sich seine Wand daher dehnt.
Der Dendrit kann (anders als bei einem „normalen“ Nerven) u.U. sehr lang sein, weil er dorthin zieht, wo der Druck gemessen werden soll.
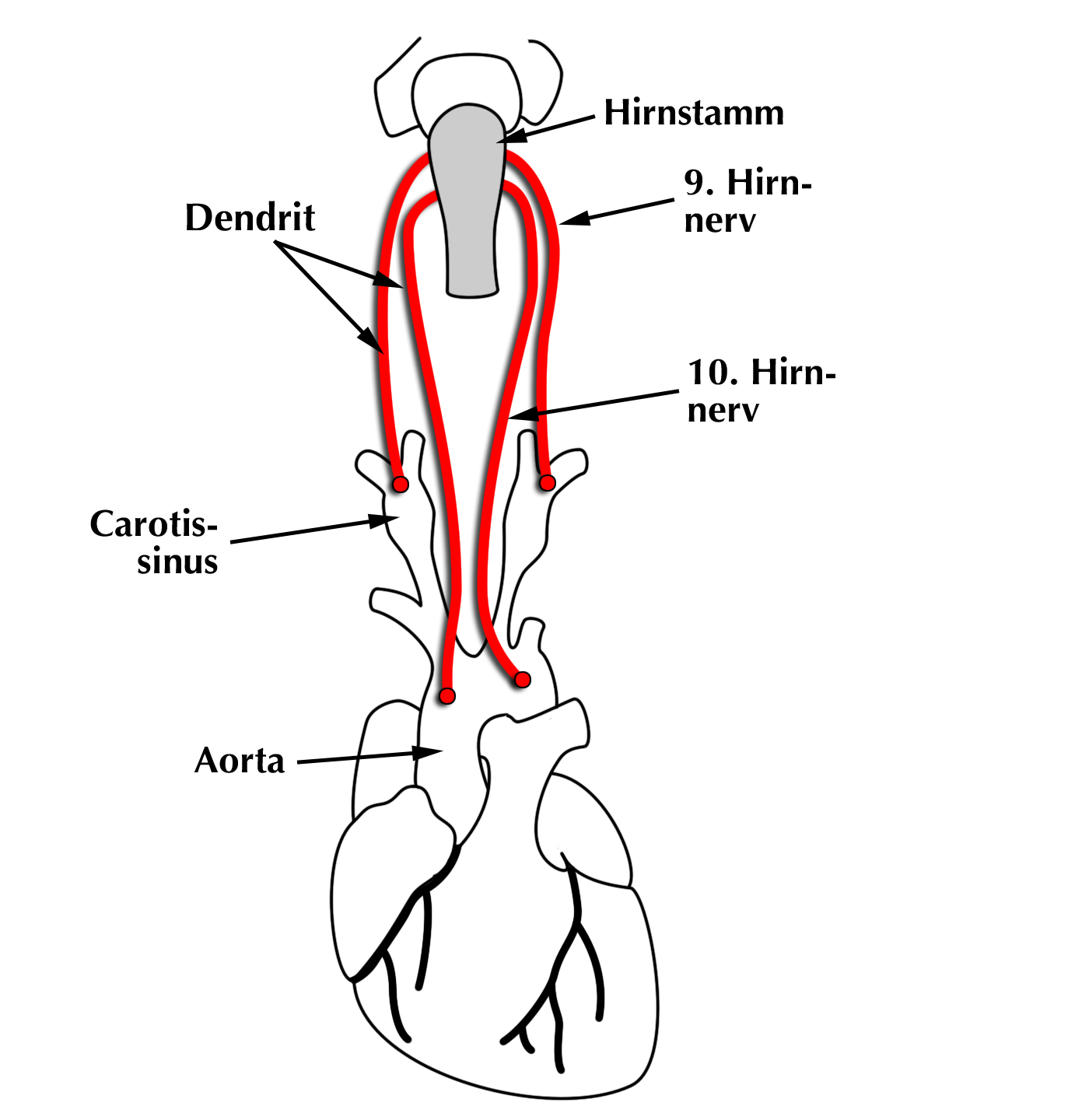 |
| Abb. 11 |
Der Ort der Druckmessung bestimmt den anatomischen Verlauf des Dendriten:
Diejenigen, deren Rezeptoren den Druck in der Aorta messen sollen verlaufen zusammen mit dem 10. Hirnnerven, die Dendriten mit den Drucksensoren für die Halsschlagader laufen zusammen mit dem 9. Hirnnerven (Abb. 11).
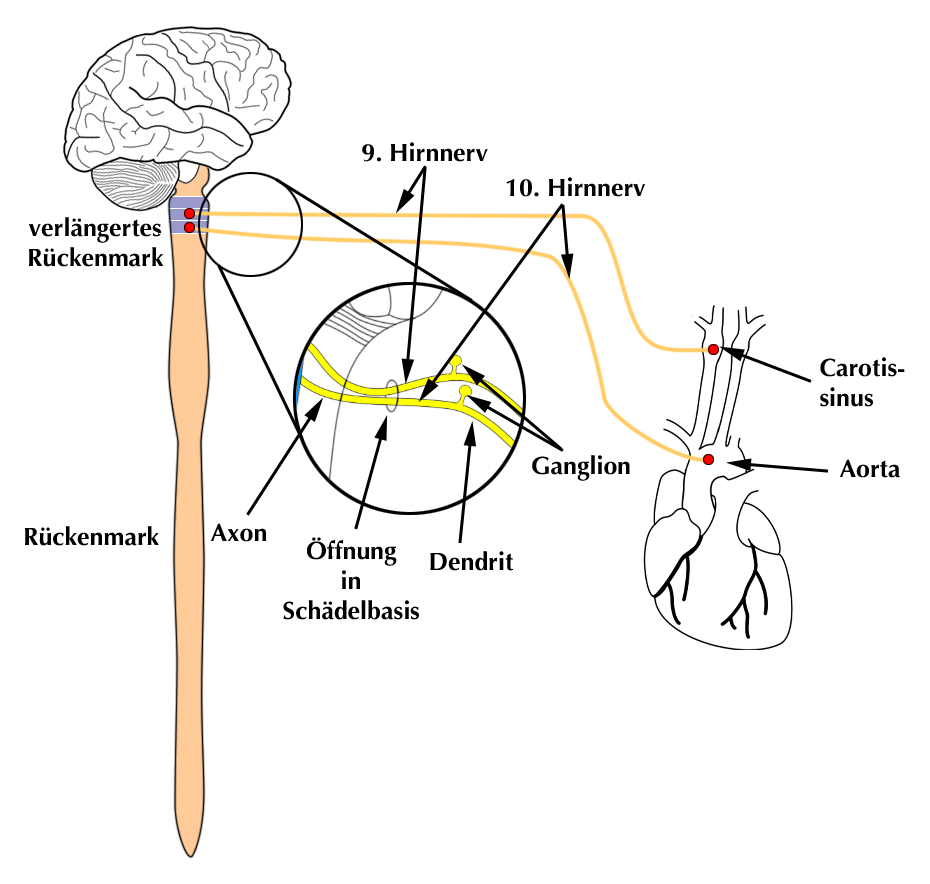 |
| Abb. 12 |
Die 2. Ausziehung des Zellkörpers der Rezeptorzelle, das Axon, zieht ins Gehirns, wo es an ein bestimmtes Kreislaufzentrum (siehe unter „Steuerzentren“) angeschlossen ist und dieses informieren kann (Abb. 12).
Druckrezeptoren befinden sich u.a. in der Haut, wo Sie Ihnen vermitteln, daß gerade Druck auf die Haut ausgeübt wird, wenn Sie Ihre Finger noch in der Tür haben, wenn sich diese gerade schließt.
Nun sind die Druckrezeptoren Ihrer Finger für die Kreislaufregularisation nicht so interessant. Es gibt aber auch Druckrezeptoren in den Wänden von Schlagadern, wo es ihre Aufgabe ist, den Blutdruck zu messen.
Sie sind die für Regulation des Kreislauf entscheidenden Sensoren. Ihre Aufgabe ist es dabei, kontinuierlich den arteriellen Blutdruck zu messen und das Kreislaufzentrum über diese Meßergebnisse zu informieren, damit dieses dafür sorgen kann, daß der Druck stets auf einem konstanten Niveau bleibt, sodaß die bedarfsgerechte Blutversorgung der einzelnen Organe gewährleistet ist. Motto: Sinkt der Blutdruck messen die Drucksensoren dies und melden es der Steuerzentrale.
Die Zentrale zieht dann aus diesen Informationen die entsprechenden Konsequenzen und wird entsprechend tätig.
Steigt der Blutdruck wird dies ebenfalls gemeldet.
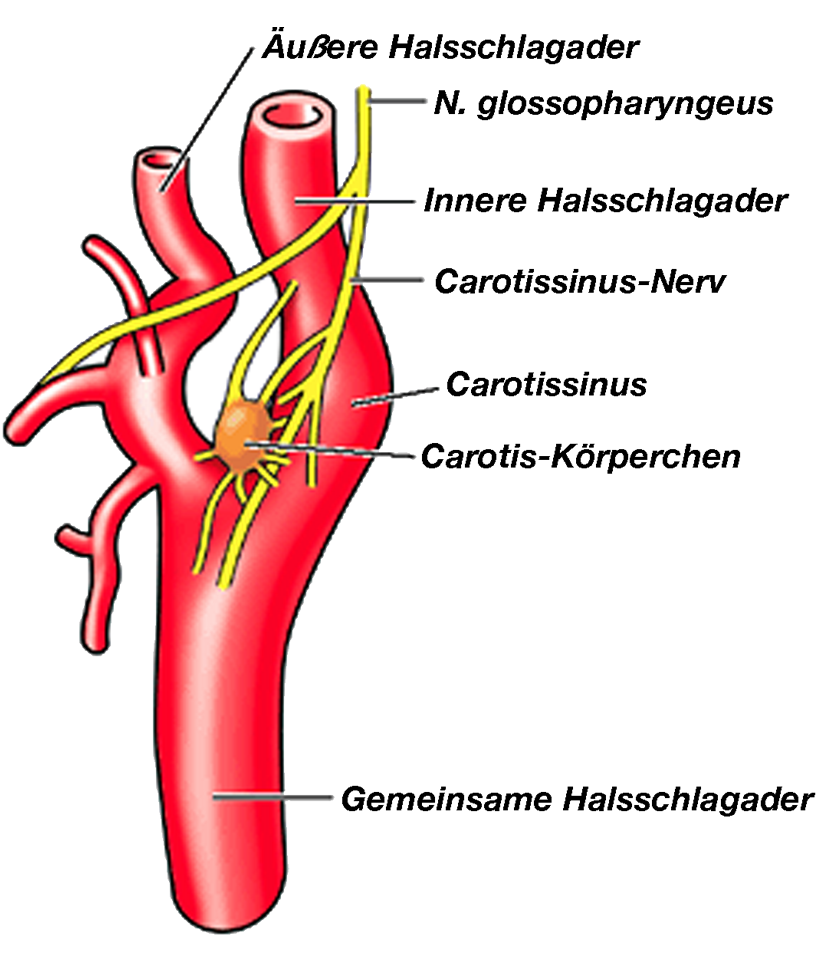 |
| Abb. 13 |
Noch ein Wort zum Carotissinus (Abb. 13):
Beim Carotissinus handelt sich um ein Ansammlung vieler Barorezeptoren, die in der Gabel der Halsschlagader (= A. carotis communis) in die äußere (= A. carotis externa) und die innere Halsschlagader (= A. carotis interna) gelegen sind. Zusammen mit den Druckrezeptoren in der Brust-Aorta ist der Carotissinus der wichtigste Druckfühler des Kreislaufes:
Mißt z.B. der Carotissinus einen zu hohen Blutdruck meldet er dies dem Kreislaufzentrum, das aus dieser Information seine Rückschlüsse zieht; meldet er einen zu niedrigen Blutdruck wird dies gemeldet und man zieht an zuständiger Stelle die entsprechenden richtigen Rückschlüsse (schrecklicher Ausdruck: „Dies wird gemeldet und man wird an zuständiger Stelle die richtigen Rückschlüsse ziehen“; klingt bedrohlich (so nach geheimer Staatspolizei oder Staatssicherheit oder KGB oder so), aber Sie wissen hoffentlich, wie ich´s meine).
Wie wichtig dieser Carotissinus ist merkt man, wenn dieser Drucksensor krank wird:
Beim „überempfindlichen Carotissinus“ reagiert er nämlich schon auf normale Drücke, z.B. den Druck auf die Haut mit dem Rasierapparat (betrifft natürlich nur Männer!) oder er reagiert dann, wenn man den Hals dreht und dabei diese Stelle des Halses unvermeidlich unter Druck setzt. In diesen Fällen meldet der überempfindliche Carotissinus dem Kreislaufzentrum nämlich: „Achtung! Massiv erhöhter Druck“.
Die Zentrale empfängt diese Meldung und kann deren Wahrheitsgehalt natürlich nicht überprüfen, sondern nimmt sie als gegeben und wahr an (wie im Leben: Das, was gemeldet wird (erst Recht wenn´s schriftlich erfolgt) muß wahr sein). Die logische Konsequenz der Zentrale besteht darin, nun alles zu unternehmen, um diesen vermeintlich massiv erhöhten Blutdruck zu senken. Dies geschieht, indem dem Herzen der Befehl gegeben wird, seine Schlagfrequenz zu verlangsamen oder sogar kurzfristig einzustellen, denn das geht am schnellsten.
Die Folge:
Kurzer Herzstillstand oder starke Verlangsamung des Herzschlags mit der Folge eines nun drastischen Blutdruckabfalls mit Schwindel oder sogar Ohnmacht.
Eigentlich ein gut gemeinter und sinnvoller Vorgang, der aber im Krankheitsfall auch zum Problem werden kann.
Dehnungsrezeptoren des Herzens
Typ: Kombinierter Rezeptor
Die Dehnungsrezeptoren des Herzens sind „kombinierte Rezeptoren“. Das bedeutet, daß sie gleichzeitig Informationen weiterleiten und ebenso ein Hormon als „Werkzeug“ bilden können.
Der Aufbau der Dehnungsrezeptoren entspricht, weil es sich um Nervenzellen handelt, prinzipiell demjenigen der Druckrezeptoren:
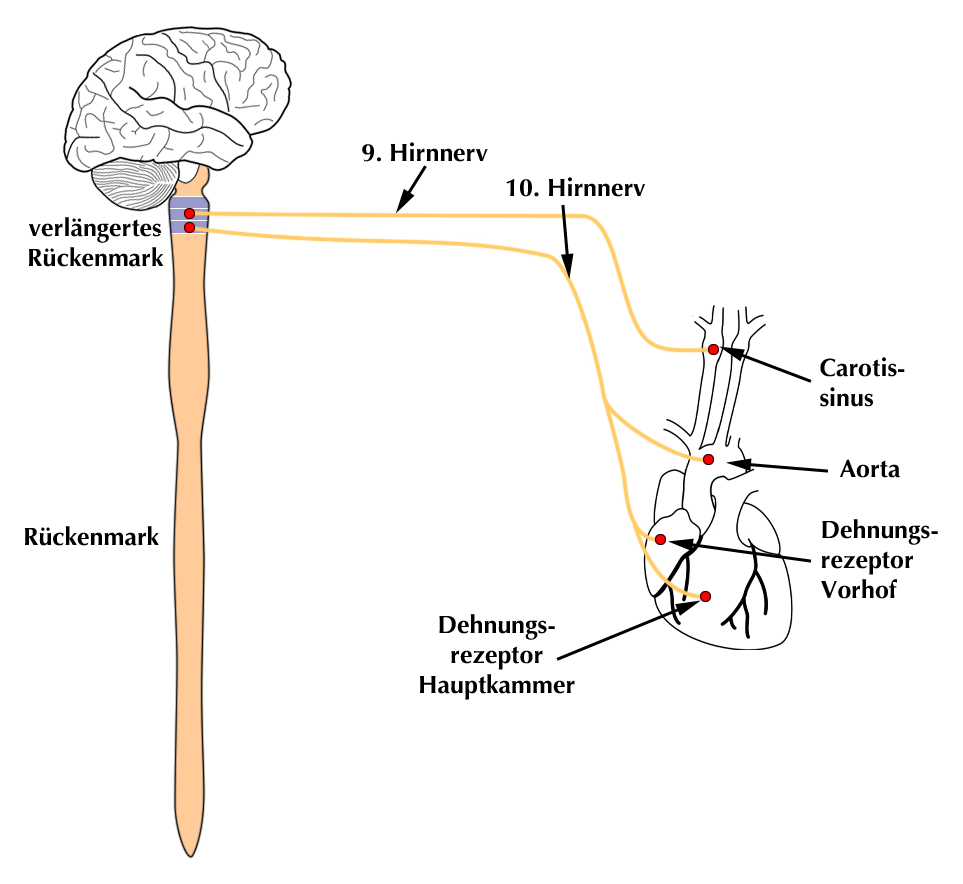 |
| Abb. 14 |
Der Zellkörper der Rezeptorzelle befindet sich im Ganglion des 10. Hirnnerven (N. vagus) im Bereich der Schädelbasis.
Die kurzen Axone der efferenten Bahn sind mit dem Kreislaufzentrum im verlängerten Rückenmark (Medulla oblongata) verbunden, über das sie ihre Informationen an ein Kreislaufzentrum im Gehirn (hypothalamisches Zentrum) schicken (Abb. 14).
Dieses Kreislaufzentrum produziert das Hormon ADH, das in der Hirnanhangsdrüse gespeichert und bei Bedarf entsprechend u.a. der Signale der Dehnungsrezeptoren ins Blut ausgeschüttet wird. Näheres über das ADH lesen Sie im Abschnitt über die „Werkzeuge“ der Kreislaufregulation, Einzelheiten über das genannte Kreislaufzentrum und die Hirnanhangsdrüse im Abschnitt über die Steuerzentralen.
Die afferente Bahn des Rezeptors in Gestalt eines langen Axon-ähnliche Dendrits zieht vom Ganglion des 10. Hirnnerven zusammen mit dem 10. Hirnnerven über den Plexus cardialis zum Herzen (siehe Abb. 14).
Hier an der Spitze des Dendrits befindet sich der eigentliche Dehnungsrezeptor.
Dehnungsrezeptoren gehören ebenso wie die Druckrezeptoren zur Gruppe der Mechanorezeptoren und sind damit spezialisierte Nervenzellen.
Streng genommen handelt es sich (siehe oben) bei den Druckrezeptoren eigentlich auch um Dehnungsrezeptoren.
Man bezeichnet aber die Rezeptoren in der Aortenwand und im Carotissinus als Druck- und die hier beschriebenen Sensoren als Dehnungsrezeptoren, obwohl beide nach demselben Prinzip arbeiten.
Dehnungsrezeptoren gibt es an verschiedenen Stellen des Körpers. In den Muskeln der Arme und Beine beispielsweise nennt man sie nach ihrem mikroskopischen Aufbau „Muskelspindeln“.
Bei diesen Muskelspindeln handelt es sich um Rezeptoren, die dafür zuständig sind, dem Gehirn mitzuteilen, die stark gedehnt ein Muskel ist, damit das Gehirn daraus ableiten kann, ob sich der Muskel für die jeweilige Situation oder beabsichtigte Tätigkeit ausreichend zusammengezogen hat oder ob er noch zu stark gedehnt ist. Solche Muskelspindel gibt es nur in der sog. Skelettmuskulatur, aber nicht am Herzmuskel.
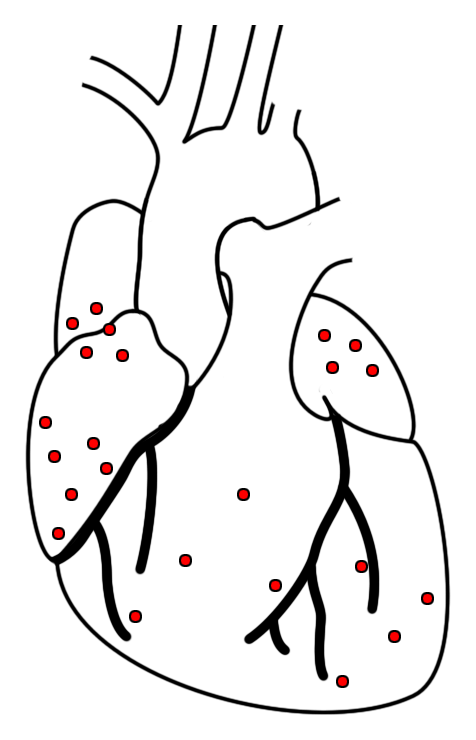 |
| Abb. 15 |
Die Dehnungsrezeptoren, die an der Regulation des Kreislaufs beteiligt sind liegen in den Muskelwänden des Herzens und zwar sowohl in den Vor- als auch in den Hauptkammern, bevorzugt in den Herzohren:
In den Vorkammern:
In größerer Ansammlung liegen sie dort, wo die Hohlvenen in den rechten bzw. die Lungenvenen in den linken Vorhof münden (Abb. 15).
Ihre Aufgabe ist es, die Dehnung und damit den Füllungszustand der Vorhöfe zu überwachen.
Enthält der Vorhof beispielsweise zu viel Blut (z.B. sammelt es sich bei einer Herzschwäche hier besonders stark an, weil es nicht in die geschwächte Hauptkammer abfließen kann) dann ist er prall gefüllt und die Vorhofwand ist daher stark gedehnt.
Enthält der Vorhof zu wenig Blut (z.B. wenn man nicht genügend trinkt) dann ist die Vorhofwand schlaff und wenig gedehnt.
Bei den Dehnungsrezeptoren der Vorkammern unterscheidet man „A“- und „B“-Rezeptoren:
- Die A-Rezeptoren messen den Dehnungszustand der Vorkammer während der Pumpphase der Vorkammer. Sie spielen bei der Kreislaufregulation kaum eine Rolle.
- Wichtiger sind die B-Rezeptoren. Sie messen die passive Dehnung der Vorhofwand während der Füllung der Vorkammer. Sie reagieren damit auf den Füllungszustand des Vorhofes.
Die Dehnungsrezeptoren sind, was ihre Informationsübertragende Funktion angeht, über Nervenfasern an das Kreislaufzentrum im Gehirn angeschlossen. Dieses Zentrum veranlaßt bei Bedarf die notwendigen Maßnahmen, indem es
- die Arbeitsweise des Herzens und
- die Weite der Blutgefäße beeinflußt.
Neben ihrer Aufgabe als Meßgerät haben die Dehnungsrezeptoren des Herzens aber auch die Möglichkeit, selbstständig zu agieren und ein bestimmtes Hormon (atriales natriuretisches Hormon (ANP), „siehe unter Werkzeuge“) zu produzieren.
Durch die Arbeitsweise als „kombinierte Rezeptoren“ können sie den „BAINBRIDGE-“ und den „GAUER-HENRY-Reflex“ auslösen, auf die ich später in diesem eBook noch eingehen werde.
In den Hauptkammern (= Ventrikel):
Die Dehnungsrezeptoren, die in die Muskelwand der Hauptkammern eingebaut sind arbeiten vom Prinzip her ebenso wie diejenigen in den Vorkammern.
Zusätzlich zur Dehnung der Herzwand und damit zur Füllung der Ventrikel können sie aber auch die Muskelkraft messen, mit der sich die Ventrikelwände während ihrer Pumpphase zusammenziehen.
Neben ihrer messenden Funktion können auch die Dehnungsrezeptoren der Ventrikelwände eigenständig das Hormon BNP ausschütten.
BNP wirkt ebenso wie das schon erwähnte ANP, was im Abschnitt über die „Werkzeuge“ noch beschrieben wird.
Die Informationen, die die Dehnungsrezeptoren der Ventrikel an die Kreislaufzentren übermitteln sind für die Regulation des Kreislaufs von großer Bedeutung, denn sie steuern vor allem die Pumpkraft des Herzmuskels. In bestimmten Situationen können die Dehnungsrezeptoren der Herzhauptkammern an dem sog. BETZOLD-JARISCH-Reflex beteiligt sein und plötzliche Ohnmachtsanfälle verursachen; Einzelheiten später später in diesem eBook.
Soviel also zu den Mechanorezeptoren.
Es gibt aber noch andere Sensoren:
Chemorezeptoren
Typ: Informativer Rezeptor
Chemorezeptoren überwachen alle chemischen Vorgänge und Zustände des Körpers, also z.B.:
- den Sauerstoffgehalt des Blutes oder der Gewebe,
- den Kohlendioxidgehalt des Blutes und bestimmter Gewebe
- den Säuregehalt des Gewebes (pH-Wert),
- den Kaliumgehalt bestimmter Flüssigkeiten und Gewebe,
- den Wasser- und den Salzgehalt des Urins und vieles mehr.
Auch bei ihnen handelt es sich um spezialisierte Nervenzellen mit Zellkörper, Axon und Dendrit, an dessen Ende sich der eigentliche Chemorezeptor befindet.
Die Chemorezeptoren sind anatomisch anders als die Druck- und Dehnungsrezeptoren aufgebaut, weil sie nämlich keinem Hirnnerven und dessen Ganglion zugeordnet sind:
Es gibt zentrale und periphere Chemorezeptoren:
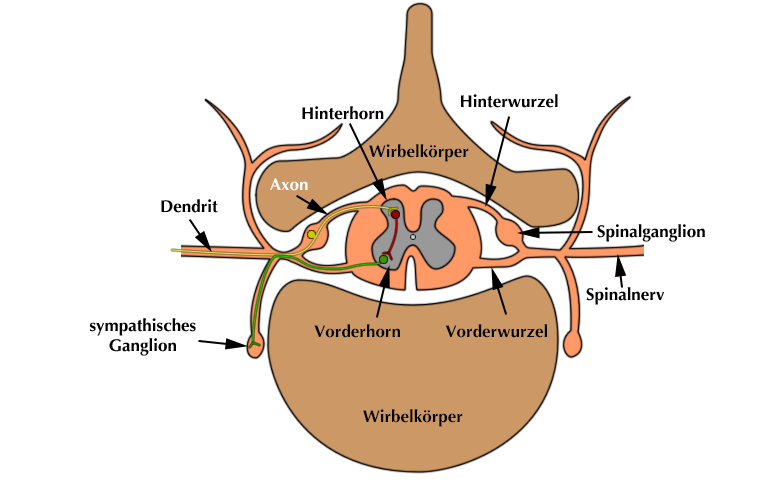 |
| Abb. 16 |
- Die zentralen Chemorezeptoren befinden sich im Gehirn, meistens in unmittelbarer örtlicher Nähe zu denjenigen Gehirnzentren, über die die verschiedenen Parameter, die gemessen werden auch gesteuert werden. Axon und Dendrit sind daher sehr kurz.
- Im Gegensatz dazu befinden sich die Zellkörper der peripheren Chemorezeptoren in unmittelbarer Nähe zur Wirbelsäule und zwar in den sog. hinteren Spinalganglien (Abb. 16).
Aus dieser Lage ergibt sich, daß ihre beiden Ausziehungen, Axon und Dendrit, anders geschaltet sind:
Der Dendrit mit dem eigentlichen Sensor verläuft aus dem Spinalganglion über den hinteren Teil der Rückenmarksnerven in den Körper an diejenige Stelle, an der etwas gemessen werden soll. Die Axone hingegen laufen in Richtung auf das Rückenmark, das sie über dessen hintere Wurzel betreten. Hier nehmen sie einen unterschiedlichen Verlauf:
- Einige Axone biegen ab und verlaufen dann nach oben ins Gehirn. Hier sind sie mit demjenigen Steuerungszentrum verbunden, das für die Regelung der jeweiligen Meßgröße zuständig ist.
- Einige andere Axone biegen jedoch nicht nach oben um, sondern verlaufen nach seitlich vorne ins Rückenmark. Hier haben sie direkten Anschluß an Zellen des sympathischen Nervensystems. Dies ermöglicht ihnen, daß die Impulse aus den Rezeptoren zu einer sofortigen Reaktion des Sympathikus im Sinne eines Reflexes führen. Ich werde hierauf im Abschnitt über die Steuerungszentren noch eingehen.
Eine erwähnenswerte Form der Chemorezeptoren sind die
Adenosinrezeptoren
Typ: Informativer Rezeptor Adenosinrezeptoren reagieren auf den Kontakt mit der chemischen Substanz Adenosin.
Sie sind nicht wie die meisten anderen Rezeptoren als spezialisierter Nerv aufgebaut, sondern sie sind in die Membran, d.h. der Hülle bestimmter Zellen eingebaut. Ihre Wirkung entfalten sie nur in diesen Zellen.
Es gibt nach heutigem Wissensstand 4 verschiedene Rezeptortypen, die sich an zahlreichen Stellen des Körpers befinden, z.B. im zentralen Nervensystem, im Herzen, in den Nieren, in Fett-, Knochen- und Immunzellen, in den Wänden bestimmter Blutgefäße oder in der Bauchspeicheldrüse. Dabei hat jeder Rezeptortyp, abhängig davon, wo er sich befindet unterschiedliche Funktionen, die sich z.T. überschneiden. Im Zusammenhang mit dem Thema dieses eBooks gibt es folgende Funktionsweisen:
- In den Herzkranzarterien verursachen sie eine Gefäßerweiterung,
- an den elektrischen Leitungsstrukturen des Herzens bewirken sie eine Verlangsamung der Erregungsüberleitung, eine Verlangsamung der elektrischen Impulsbildung und der Überleitung der elektrischen Impulse von den Vor- in die Hauptkammern und
- in den Nieren verursachen sie eine Verengung derjenigen kleinsten Blutgefäße, die das Blut in die einzelnen Filtereinheiten der Nieren (d.i. die Glomeruli, siehe unter Niere im Abschnitt über die Werkzeuge).
Die Adenosinrezeptoren des Herzens spielen unter normalen Umständen keine Rolle, wenngleich sie diagnostisch oder therapeutisch bei bestimmten Erkrankung und Untersuchungen eine wichtige Bedeutung haben.
Adenosinrezeptoren sind aber die Grundlage für den sog. tubuloglomerulären Reflex, der bei der Regelung der Filterfunktion der Nieren bedeutsam ist (siehe etwas später beim juxtaglomerulären Apparat).
Osmorezeptoren
Typ: Informativer Rezeptor
Auch bei ihnen handelt es sich um spezialisierte Nervenzellen. Deren Sensor am Ende des Dendriten kann die Konzentration gelöster osmotisch wirksamer Moleküle außerhalb von Zellen messen.
Osmotisch wirksame Moleküle sind solche Stoffe, die an der „Osmose“ beteiligt sind und sie verursachen:
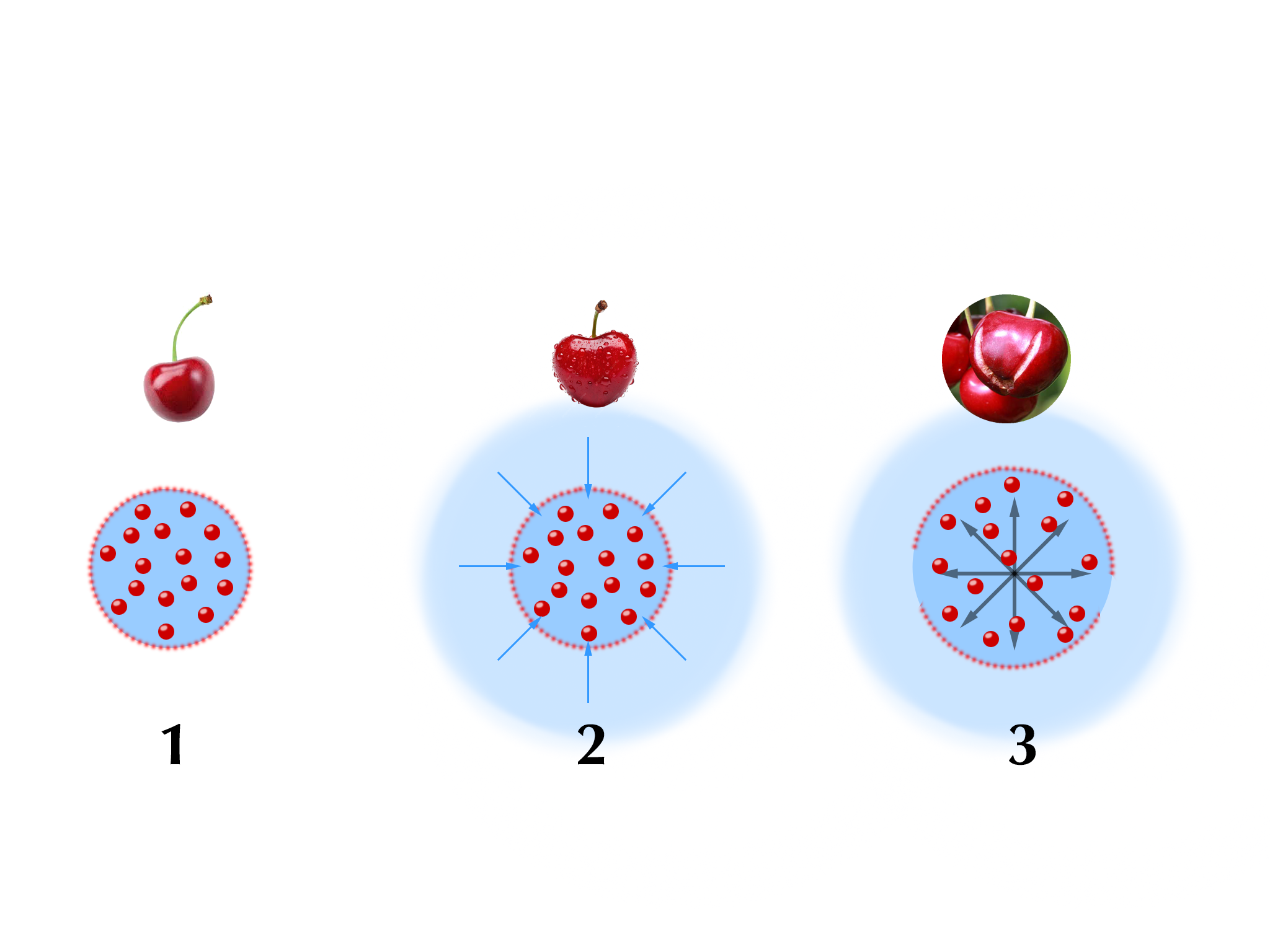 |
| Abb. 17 |
Als Osmose bezeichnet man den Fluß von Teilchen durch eine teildurchlässige Trennschicht. Als anschauliches Beispiel hierfür eine Kirsche bei Kontakt mit Regenwasser: Im Inneren einer Kirsche befinden sich viel Zucker (= hohe osmotische Konzentration oder hohe Osmolarität) und zahlreiche andere gelöste Teilchen.
Umgeben ist die Kirsche von einer Haut, die sehr wohl für Wasser, nicht aber für Zucker durchlässig ist (= teildurchlässige Trennschicht) („1“ in Abb. 17).
Wird die Kirsche bei Regen naß dann befindet sich auf der äußeren Seite der Kirschhaut das Regenwasser, das keinen Zucker enthält und auf der inneren Seite der Haut das Kirschfleisch mit dem vielen Zucker, es besteht somit ein Konzentrationsunterschied für den Zucker („2“ in Abb. 17).
Nun ist es ein Naturgesetz, daß in solchen Situationen ein Ausgleich der Konzentrationen auf beiden Seiten der Trennwand erfolgt. Der Zucker kann das Kirschfleisch nicht verlassen, weil die Kirschhaut dies verhindert. Die Haut ist aber für das Wasser durchlässig. Also wird das Regenwasser durch die Haut ins Kirschinnere eindringen, um die dort hohe Zuckerkonzentration zu verdünnen und am Ende dafür zu sorgen, daß auf beiden Seiten der Haut dieselbe Zuckerkonzentration herrscht („3“ in Abb. 17).
Durch das eindringende Wasser quillt die Kirsche auf bis der Druck im Kirschinneren so hoch ist, daß die Kirsche platzt. In diesem Beispiel ist der Zucker die osmotisch wirksame Substanz, den gesamten Vorgang mit der Wanderung des Wassers bezeichnet man als Diffusion.
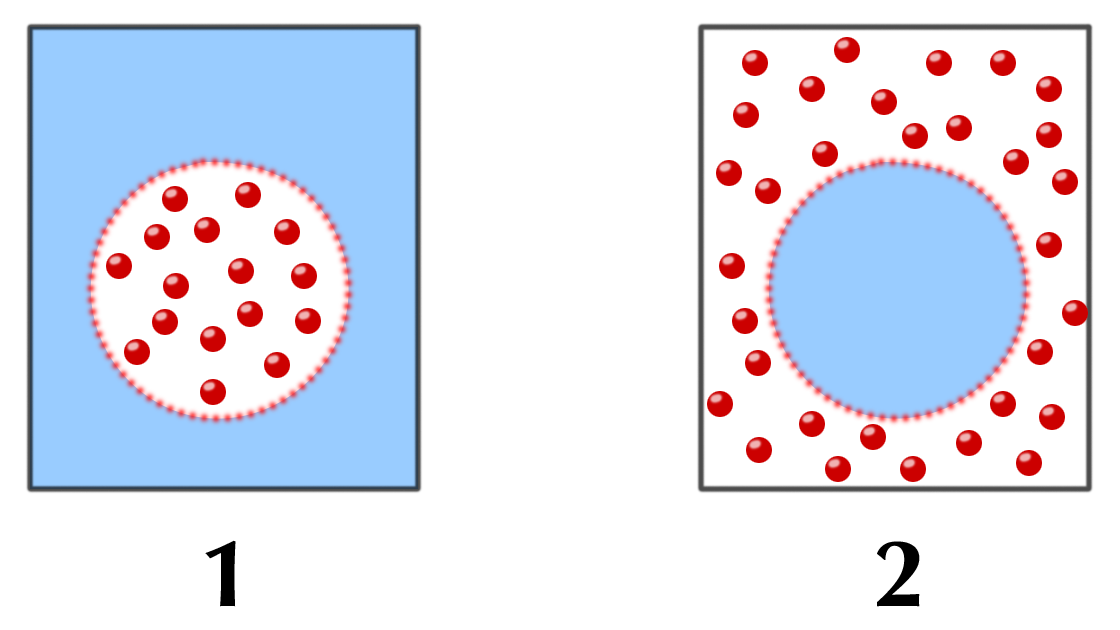 |
| Abb. 18 |
Diesen Vorgang machen sich die Osmorezeptoren zu Nutze, wobei sie im Sinne des oben beschriebenen Beispiels als Kirsche fungiert:
Ist die osmotische Konzentration (= Osmolarität) außerhalb des Osmorezeptors, d.h. im Blut, geringer als die Osmolarität innerhalb des Rezeptors („1“ in Abb. 18) bedeutet dies, daß sich außerhalb der Rezeptorzelle zuviel oder (besser gesagt:) daß sich innerhalb der Zelle zu wenig Wasser befindet. Dies bedeutet, daß nach dem Gesetz der Diffusion Wasser in die Zelle eindringt und „sie aufbläht“.
Auf eine solche „Aufblähung“ reagieren die Osmorezeptoren mit einer Vermehrung Ihrer Signale.
Ist die osmotische Konzentration außerhalb des Osmorezeptors, d.h. im Blut, andererseits höher als innerhalb des Rezeptors (= außerhalb der Zelle zu wenig, innerhalb der Zelle zuviel Wasser) („2“ in Abb. 18) verläßt Wasser nach dem Gesetz der Diffusion die Zelle, d.h. sie verkleinert sich und schrumpft. Auf eine solche „Schrumpfung“ reagieren die Osmorezeptoren mit einer Verminderung Ihrer Signale.
Osmorezeptoren gibt es hauptsächlich an 2 Stellen im Körper:
In einem der Kreislaufzentren im Gehirn und in den Nieren.
In den Nieren ist dies der „juxtaglomeruläre Apparat“, über den Sie etwas später mehr lesen werden.
Und an der Regelung der Osmolarität sind 3 Werkzeuge beteiligt: Das Durstgefühl, das Hormon ADH und die Hormonvorstufe Renin.
Die 3 o.g. Mechanismen wirken dabei wie folgt:
- Über das Durstgefühl wird der Wasserhaushalt des Körpers gesteuert. Bei Durst trinkt das Lebewesen mehr und der Wassergehalt des Blutes nimmt zu, seine Osmolarität nimmt ab.
- Das ADH (Einzelheiten unter „Werkzeuge“) fördert die Rückgewinnung von Wasser in den Nieren. Die Urinmenge nimmt ab, der Wassergehalt des Blutes nimmt zu, seine Osmolarität ab.
- Das Renin (Einzelheiten siehe unter „Werkzeuge“) ist eine Hormonvorstufe, die zur vermehrten Bildung u.a. von Aldosteron (Einzelheiten ebenfalls unter „Werkzeuge“) führt.
- Aldosteron wiederum ist ebenfalls ein Hormon, daß die Rückgewinnung von Wasser in den Nieren fördert.
Alle diese Mechanismen führen dazu, daß, je nach Erfordernis mehr oder weniger Wasser ins Blut aufgenommen wird. Wird mehr Wasser aufgenommen kommt es zu einer „Verdünnung“ der osmotisch wirksamen Teilchen im Blut, dessen osmotische Konzentration (d.i. die Osmolarität) dadurch abnimmt. Wird weniger Wasser aufgenommen werden die Teilchen „eingedickt“, d.h. die Osmolarität nimmt zu. •
Der Regelmechanismus funktioniert folgendermaßen:
Osmolarität im Blut zu hoch:
In diesen Fällen gibt es im Blut zuviele osmotisch wirksame Teilchen oder anders herum: Es fehlt also Wasser, die Osmolarität des Blutes ist zu hoch.
- In den Nieren führt dies zur vermehrten Ausschüttung von Renin.
- Das Renin führt zur vermehrten Bildung u.a. von Aldosteron.
- Vermehrtes Aldosteron bewirkt in den Nieren eine vermehrte Rückgewinnung von Wasser ins Blut.
- Dieses Wasser verdünnt die Teilchen im Blut, sodaß die Osmolarität des Blutes abnimmt.
- Das Renin aktiviert aber zusätzlich im Gehirn ein bestimmtes Kreislaufzentrum.
- Dieses Kreislaufzentrum aktiviert die Hirnanhangsdrüse, die nun mehr ADH ausschüttet.
- Das ADH fördert an der Niere eine verstärkten Wasserrückgewinnung, d.h. es wird vermehrt Wasser ins Blut aufgenommen und die ursprünglich erhöhte Osmolarität des Blutes sinkt wieder ab.
- Ein anderes Steuerzentrum im Gehirn (Nucleus supraopticus des Hypothalamus) löst ein Durstgefühl aus.
- Dieses veranlaßt den Körper, mehr zu trinken. Damit wird über den Magen-Darm-Trakt vermehrt Wasser ins Blut aufgenommen, was dort ebenfalls zur Senkung des Osmolarität führt.
Osmolarität im Blut zu gering: Hier laufen die Mechanismen genau anders herum:
- In den Nieren wird weniger Renin ausgeschüttet, was auch zu einer verminderten Bildung von Aldosteron führt. Hierdurch wird weniger Wasser zurückgewonnen und mehr ausgeschieden, die Osmolarität des Blutes nimmt zu.
- Das Kreislaufzentrum im Gehirn schüttet weniger ADH aus. In der Folge wird mehr Wasser ausgeschieden und die Osmolarität des Blutes steigt.
- Auch das Durstgefühl nimmt ab, sodaß das Lebewesen weniger Wasser trinkt, was die Osmolarität des Blutes ebenfalls erhöht.
Neben dem juxtaglomerulären Apparat der Nieren und dem Gehirn finden sich Osmorezeptoren auch im Rachen, im Verdauungstrakt und in der Leber. Um welche Zellen es sich hierbei handelt, wohin die Impulse dieser Zellen übertragen werden und was sie bewirken ist allerdings nicht genau bekannt.
Thermorezeptoren
Typ: Informativer Rezeptor Thermorezeptoren sind Sensoren, die auf Wärme reagieren.
Sie werden in diesem Zusammenhang nur deshalb besprochen, weil die Körpertemperatur auch Auswirkungen auf die Regulation des Kreislaufes hat.
Prinzipiell gibt es 2 Ort, an denen sich Thermorezeptoren befinden: Im Gehirn (= zentrale Thermorezeptoren) und an anderen Stellen des Körpers (= periphere Rezeptoren).
- Die zentralen Rezeptoren finden sich in einer bestimmten Gegend des Gehirn, nämlich im Hypothalamus. Ihre Zellkörper befinden sich in unmittelbarer Nähe zum Steuerungszentrum der Wärmeregulation. Die Dendriten mit den eigentlichen Wärmesensoren messen die Temperatur im Gehirn und geben diese Information über das Axon an das Wärmeregulationszentrum weiter.
- Die peripheren Thermorezeptoren befinden sich in der Haut und den Schleimhäuten der Eingeweide.
Weil es sich ebenso wie die anderen Rezeptoren um spezialisierte Nervenzellen handelt entspricht auch der Aufbau der Rezeptoren demjenigen einer Nervenzelle:
Ihre Zellkörper liegen in den Spinalganglien bzw. in den Ganglien von Hirnnerven. Von hier aus senden sie ihre teilweise sehr langen Dendriten in die Haut, wo ihre freien Enden mit den Wärmesensoren nahe der Hautoberfläche liegen. Sie erzeugen elektrische Impulse, deren Frequenz sich in Abhängigkeit von der Temperatur verändern.
Der andere Ausläufer der Zellen, das Axon, läuft zum Rückenmark. Hier im Hinterhorn werden seine Impulse entweder auf andere Nervenzellen umgeschaltet, die die Impulse bis zum Gehirn und dem hier befindlichen Wärmeregulationszentrum im Hypothalamus weiterleiten. Oder sie verlaufen innerhalb des Rückenmarks zu dessen Seitenhorn, wo sie an das sympathische Nervensystem angeschlossen sind und dessen sofortige Reaktion verursachen (siehe etwas später im Abschnitt über die Steuerungszentren unter dem Punkt „lokale Zentren“). Entweder über das Steuerungszentrum im Gehirn oder über die lokalen Zentren im Rückenmark werden dann ggfs. Maßnahmen veranlaßt, die die Körpertemperatur wieder normalisieren sollen, z.B. bei Aufhitzung des Körpers im Sommer:
- Die Hautgefäße werden erweitert, damit mehr Wärme in die Haut gelangt, von wo aus sie dann abgestrahlt wird (bei Kälte verengen sich die Hautgefäße),
- es wird die Produktion von Schweiß angeregt, der den Körper mit Hilfe seiner Verdunstung abkühlen soll und schließlich
- wird das Herz zum schnelleren und kräftigeren Schlagen angeregt, damit die erweiterten Hautgefäße stets gut mit Blut versorgt werden.
Juxtaglomerulärer Apparat
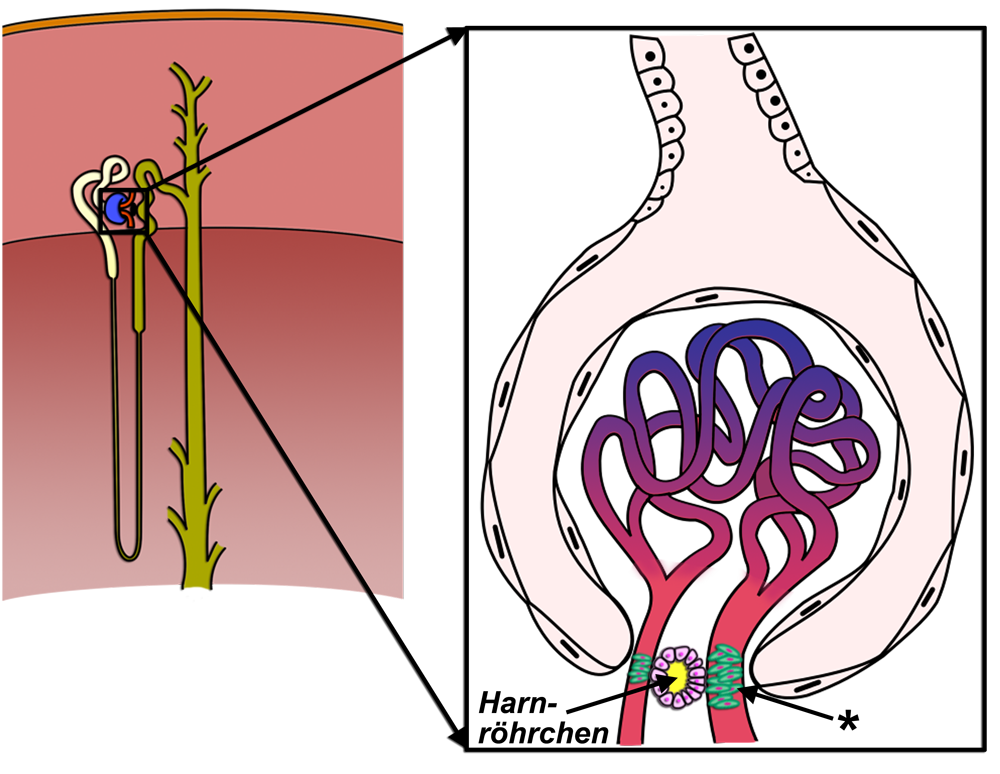 |
| Abb. 19 |
| * = juxtaglomerulärer Apparat |
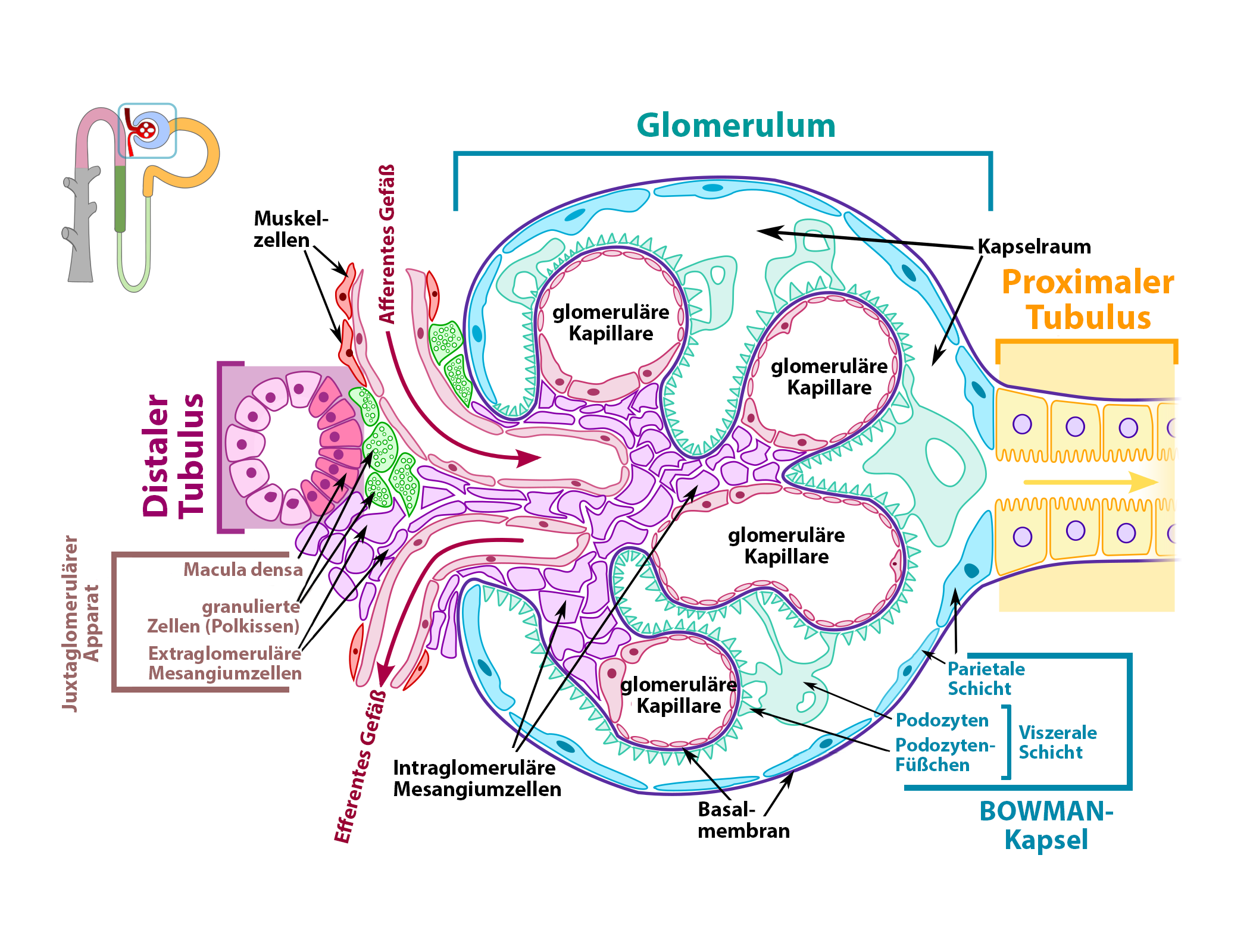 |
| Abb. 20 |
| Nach: M. Komorniczak, Wikipedia |
Typ: Hormonproduzierender Rezeptor
Der juxtaglomeruläre Apparat liegt in den Nieren (siehe unter „Werkzeuge“) unmittelbarer Nähe des Eingangs in die BOWMAN-Kapsel in engem Kontakt mit den zu- und abführenden Gefäßen und einem Teil des Harnröhrchens (Abb. 19).
Über die Arbeitsweise und den Aufbau der Nieren werden Sie später im Abschnitt über die Werkzeuge Genaueres lesen.
Es handelt sich beim juxtaglomerulären Apparat um eine Ansammlung von 3 Arten spezialisierter Zellen. Dabei handelt es sich um die sog. Macula densa, um die granulierten Zellen und um die extraglomerulären Mesangiumzellen (Abb. 20).
- Macula densa: Es handelt sich um Osmorezeptoren, die sich in der Wand des aufsteigenden Harnröhrchens (= distaler Tubulus, Einzelheiten im Abschnitt über die Niere unter den Werkzeugen) befinden. Diese Osmorezeptoren messen den Salzgehalt und damit die Osmolarität des Urins. In bestimmten Situationen schütten Sie das Hormon „Adenosin“ aus. Dieses Adenosin tritt dann in Kontakt mit den Adenosin-Rezeptoren in der Muskelwand der aus den Nierenkörperchen austretenden Blutgefäße und führt zu deren Verengung. Mit diesem Mechanismus wird die Menge des in der Niere gebildeten Harn gesteuert. Mehr zu diesem „tubuloglomerulären Reflex“ erfahren Sie später in diesem eBook unter den Regelkreisen. Die Macula densa ist wegen ihrer Meßfunktion mit Hilfe der Osmorezeptoren und der gleichzeitigem hormonproduzierenden Funktion ein kombinierter Rezeptor.
- Granulierte Zellen: In diesen Zellen, die man auch als Polkissen bezeichnet, wird das Hormon Renin gebildet, das bei der Steuerung von Blutdruck und Flüssigkeitsmenge im Kreislauf eine sehr wichtige Rolle spielt. Die Zellen des Polkissens sind der hormonproduzierende Teil des juxtaglomerulären Apparats.
Die Ausschüttung des Renins wird durch die Chemorezeptoren der Macula densa in Abhängigkeit vom Natrium- und Chloridgehalt des Harns ausgelöst. Genaueres über das Renin erfahren Sie etwas später im Abschnitt über die Werkzeuge, über die Steuerung seiner Freisetzung und seine Auswirkungen im Abschnitt über die Regelkreise.
Renin wird in den folgenden Situationen freigesetzt:
- Bei vermindertem Blutdruck (gemessen durch die Blutdrucksensoren (Barorezeptoren) in den zuführenden Gefäße der Nierenkörperchen und
- bei erniedrigte Konzentration von Kochsalz im Harn (gemessen durch die Osmorezeptoren in den Harnröhrchen). Neben seiner Funktion als Rezeptor ist der juxtaglomeruläre Apparat aber zusätzlich auch noch direkt an das sympathische Nervensystem angeschlossen. Über diese Verbindung können sympathischen Kreislaufzentren im Gehirn, wenn es notwendig ist, den juxtaglomerulären Apparat und damit die Ausschüttung von Renin direkt beeinflussen (auf das sympathische Nervensystem werde ich im nächsten Abschnitt über die „Kommunikationswege“ noch genauer eingehen).
- Extraglomeruläre Mesangiumzellen. Sie haben bei der Regulation des Kreislaufes keine Aufgaben. Dennoch sind sie wichtig für die Funktion der kleinen Blutgefäßchen im Inneren der BOWMAN-Kapsel (= glomeruläre Kapillaren), weil sie sich zusammenziehen und damit den Blutfluß in diesen Kapillaren steuern können.
Soviel zu den wichtigsten Sensoren, die Einfluß nehmen auf die Regulation des Kreislaufes. Kommen wir nun zu den Werkzeugen der Kreislaufregulation.
Werkzeuge
Es gibt verschiedene Werkzeuge, mit denen der Kreislauf beeinflußt und reguliert werden kann:
Schließmuskel der Schlagadern
Es handelt sich um Muskeln, die den Zufluß zu einzelnen Organen oder Körpergebieten regeln.
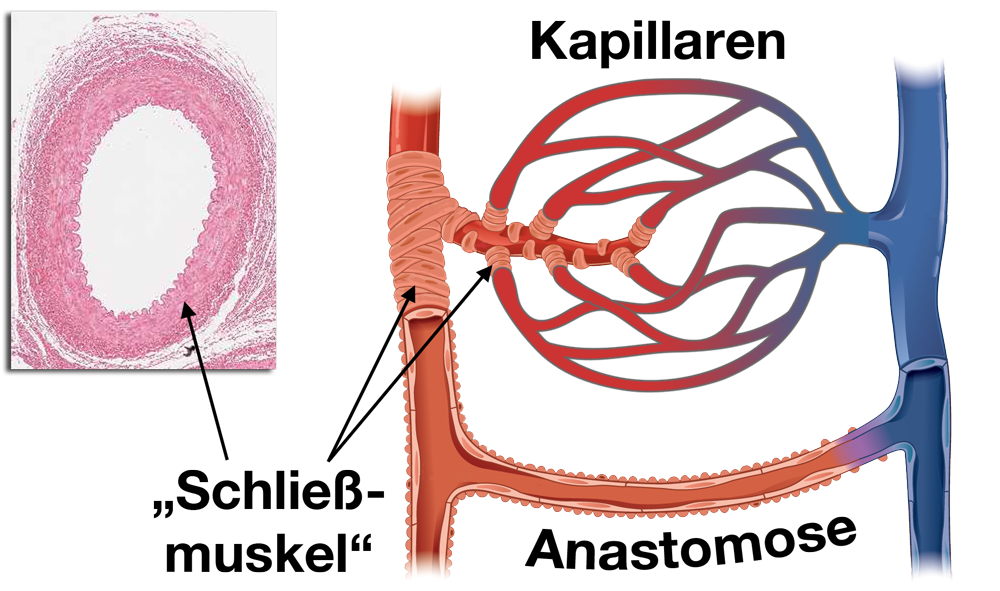 |
| Abb. 21 |
Sie funktionieren wie die Regler an den Heizkörpern:
Dreht man sie auf fließt viel Wasser in den Heizkörper, dreht man sie zu fließt entsprechend weniger. Die Verschlußmuskeln der Arterien arbeiten ebenso:
Es handelt sich um Fasern glatten Muskels, die in der Muskelschicht der Gefäßwand liegen und den Innenraum der kleinen Arterie umschlingen („Schließmuskel“ in Abb. 21).
Ziehen sich diese Muskelfasern zusammen verengen Sie Innenraum des Gefäßes, sodaß sein Durchmesser kleiner wird und weniger Blut hindurch strömen kann. Erschlaffen Sie wird der Durchmesser des Gefäßes wieder weiter und es kann mehr Blut hindurch strömen.
Die Steuerung dieser Schließmuskeln erfolgt dadurch, daß der Körper durch verschiedene Meßfühler bemerkt, daß in einer bestimmten Gegend wenig oder viel Blut benötigt wird. Wird wenig Blut benötigt verengen die Schließmuskeln das Gefäß, wird viel Blut benötigt erweitern sich die Gefäße.
Arteriovenöse Anastomosen
|
| Abb. 22 |
Bei diesen Gefäßen handelt es sich um Gefäße, die das Kapillarnetz umgehen, sozusagen eine Umgehungsstraße (Abb. 22).
Sie leiten Blut um ein Gebiet herum, das gerade wenig Blut benötigt. Auf diese Weise wird das umgangene Kapillarsystem nicht mehr durchblutet und das viele Blut, das ansonsten durch die Kapillaren flösse wird nun direkt aus den Schlagadern in die Vene geleitet, sodaß es von hier aus an diejenigen Stellen transportiert werden kann, die einen dringenderen Blutbedarf haben.
In den Wänden dieser Anastomosen befinden sich ebenfalls Schließmuskeln, die in der Lage sind, diese Umgehungsgefäße zu öffnen oder zu schließen, gesteuert wird dieses System ebenfalls über die Messung des Blut- und Sauerstoffbedarfes des Gewebes, die von den Meßfühlern vorgenommen wird. Das klassische Beispiel für solche Anastomosen ist der sogenannte „von Euler-Liljestrand-Reflex“ der Lungen, den ich unter den Autoregulationsmechanismen auf der lokalen Ebene noch eingehen werde.
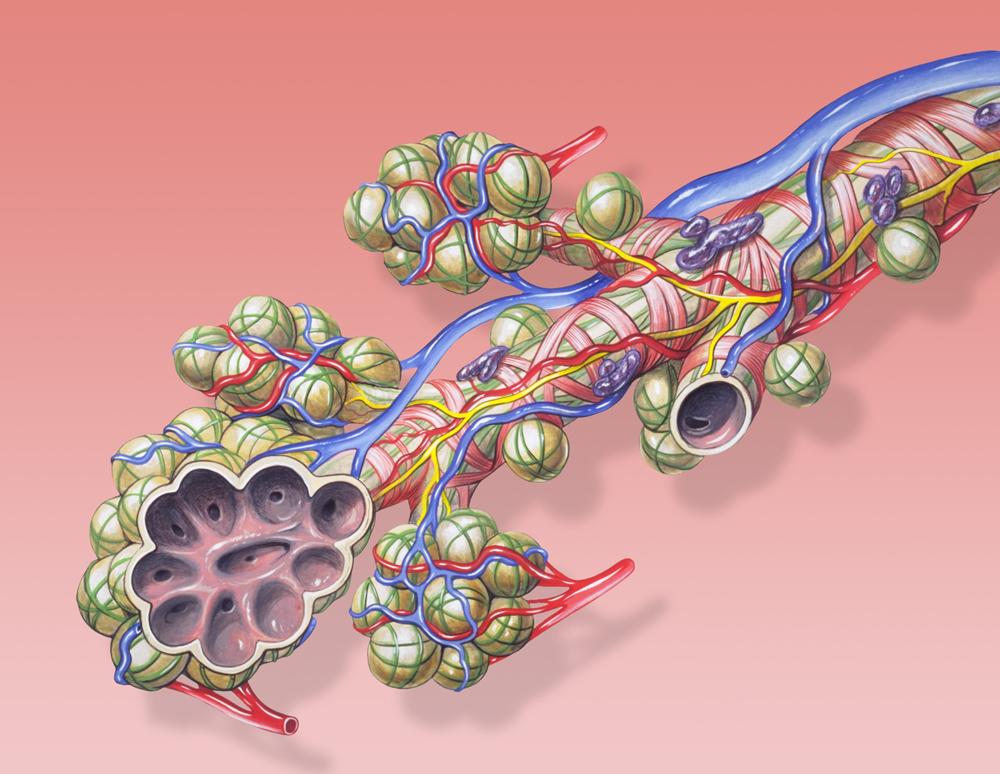 |
| Abb. 23 |
| Diese Abbildung stammt von David Lynch, einem genialen medizinischen Illustrator. Das Bild ist im Internet frei erhältlich. |
Der Reflex regelt die Durchblutung der Lunge in Abhängigkeit von deren Belüftung:
Werden bestimmte Lungenanteile vermindert belüftet wird auch die Durchblutung dieser Lungenanteile automatisch vermindert (Abb. 23).
Diese Durchblutungsverminderung erfolgt dadurch, daß sich arteriovenöse Anastomosen öffnen, die das Blut an den jeweiligen Lungenteilen vorbei leiten.
Ein weiteres Werkzeug der Kreislaufregularisation ist die
Weite der Körpervenen
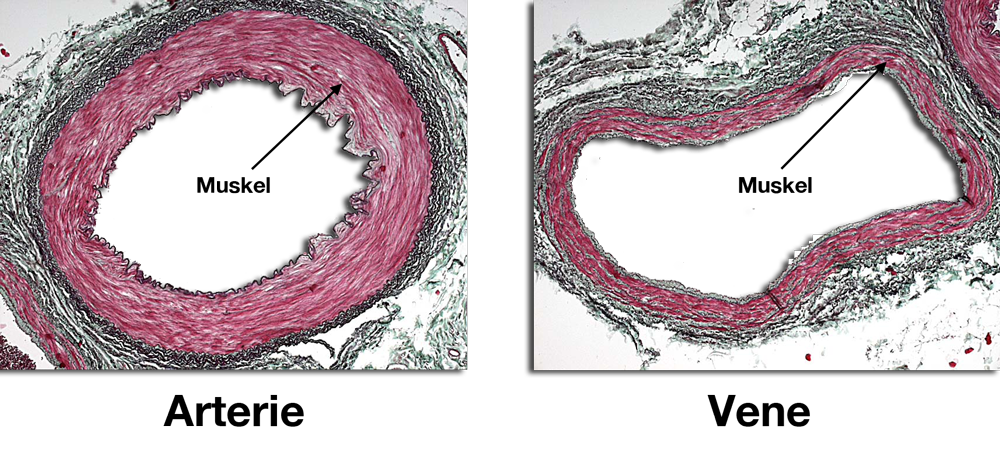 |
| Abb. 24 |
Wenn Sie sich an das System des Kreislaufes erinnern dann werden Sie wissen, daß die Arterien die Aufgabe haben, daß sauerstoffreiche Blut zu den einzelnen Organen zu transportieren und die Venen das verbrauchte Blut wieder zurück zu leiten. Dabei haben die Venen aber nicht nur die Aufgabe eines „Abwasserkanals“, sondern sie dienen auch noch als Blutspeicher.
Wenn Sie sich einmal die Abb. 24 ansehen erkennen Sie sofort, daß die Arterien eine sehr dicke und die Venen eine viel dünnere muskulöse Wand haben. Das hat etwas mit der Funktion der Gefäße und vor allem mit dem Druck zu tun, der in den jeweiligen Gefäßen herrscht (Arterie = hoher Blutdruck, Vene = niedriger Druck).
Sind die Muskeln der Venenwand straff gespannt dann ist der Innenraum der Gefäße eng. Und das bedeutet, daß das Blut, das gerade aus den Kapillaren gekommen ist schnell zum Herzen und zur Lunge weitergeleitet wird.
Sind die Muskelwände der Venen aber schaff dann erweitert sich der Innenraum der Venen durch den hier herrschenden (wenn auch geringen) Druck. Enge Venen können nicht viel Blut speichern, weite Venen aber eine ganze Menge.
Das Ganze funktioniert wie das Vorflutgelände eines Flusses:
Bei normalem Wasserstand ist das Gelände nicht geflutet und alles Wasser befindet sich im Fluß. Bei Hochwasser hingegen geben die Deiche an bestimmten Stellen nach oder es werden Flutventile geöffnet. Ein großer Teil des Wasser strömt nun ins Vorflutgelände, das hierdurch als eine Art Wasserspeicher dient und Städte und wichtige Straßen vor der Überschwemmung bewahren soll. Genauso funktioniert das mit den Venen auch:
Benötigt der Körper gerade viel Blut sind die Venen eng, das Blut wird (wie gesagt) schnell zum Herzen und den Lungen weitergeleitet und dann erneut in den Körper gepumpt. Benötigt der Körper aber gerade weniger Blut dann erweitern sich die Venen und die gerade nicht benötigte Blutmenge „versickert“ hier und liegt in Reserve. Wird dann wieder mehr Blut benötigt verengen sich die Venen wieder und führen das „gespeicherte“ Blut dadurch wieder dem Kreislauf zu.
Etwa 70-80% des gesamten Blutes eines Menschen, also etwa 4-5 l befindet sich in den Venen und wenn sich alle Venen des Körpers schlagartig erweitern würden käme praktisch kaum noch Blut am Herzen an und der Blutdruck würde ins Bodenlose abfallen. Damit dies nicht geschieht muß die Weite der Venen immer der jeweiligen Situation angepaßt sein.
Dabei erfolgt die Steuerung der Weite der Venen anhand des Blutdrucks, der an verschiedenen Stellen gemessen wird. Die Informationen der Barorezeptoren werden an die entsprechenden Kreislaufzentren übermittelt, die dann darüber entscheiden, ob Blut im Kreislauf benötigt wird (Venen werden verengt) oder ob ihm etwas Blut entzogen werden kann (Venen werden erweitert, sodaß mehr Blut in den Venen „versackt“).
Die Weite der Venen wird durch den Sympathikus (verengt die Gefäße) und den Parasympathikus (erweitert sie) gesteuert.
Soviel zu den lokalen Werkzeugen der Kreislaufregularisation. Weil Sie hier aber ein Buch über die Funktion des Herzens vor sich haben ist es eigentlich klar, daß auch das Herz selber bei der Kreislaufregularisation mitspielt.
Das Herz
Das Herz greift in die Regulation des Kreislaufes mit 3 Mechanismen ein:
- Den STARLING-Mechanismus
- der Inotropie und
- der Herzfrequenz
STARLING-Mechanismus
Der Herzmuskel und seine Pumpkraft ist zwar derjenige Teil des Körpers, der den Kreislauf in Gang hält, dennoch kann auch er auf verschiedenen Gegebenheiten und den sich hieraus ergebenden Notwendigkeiten reagieren, weshalb die Pumpkraft des Herzens variabel ist.
Für diese eigene Regularisation seiner Pumpkraft ...........